運動

脳幹の電気刺激により身体を意のままに操作し、心臓や肺を停止させ、様々な生理現象を引き起こすことができます。
それ以外の脳領域でも脳幹ほどの効力はないものの、似たような現象を引き出すことができます。
これを悪用すると、直接的には心停止や呼吸停止などを引き起こし、間接的には身体を操作して事故などを引き起こし、証拠を残さずに個人を暗殺することさえ可能になると思われます。
目次All_Pages
回路
脳幹
運動を制御する神経回路は、神経系全体に分布しています。(Ruder and Arber 2019)
運動皮質や大脳基底核などは運動の計画に関与する上位領域で、脊髄は最終的に身体の動作を実行する下位領域です。
脳幹はこの2つの領域の間に位置する重要なインターフェースで、多様な運動行動の制御に重要な神経回路を分散して擁しています。
基本的運動は脳幹以下の機能だけで実現できることが知られています。
前脳がなく脳幹が健全なカエルは、跳躍、足踏み、水泳など、正常なカエルが示すほとんどの行動を実行します。(Roh et al. 2011)
前脳がなく脳幹が健全な幼児 (水無脳症) は、正常な幼児と同様、泣き、笑い、乳をのみ、眼、顔、手足を動かします。(Saper et al. 2013)
運動モジュール
脊椎動物の運動システムは、少数の運動モジュールを組み合わせることで多様な運動を生み出すことができると考えられています。(Bizzi et al. 2008)
運動モジュールとは筋シナジーとも呼ばれ、特定の筋群を特定のパターンで活性化することで特定の運動出力を生み出す、脊髄に存在する機能単位です。
例えばカエルの防御キックは3つの運動モジュールから構成され、それぞれのモジュールに割りあてる振幅と遅延時間を変更することで、多様なキックのパターンを生み出すことができます。(d’Avella et al. 2003)
脳幹がなければこれらの運動モジュールは機能せず、脳幹は個別の運動モジュールを制御する領域であると考えられています。 (Roh et al. 2011, Ruder and Arber 2019)
実際、脳幹を電気刺激すると、四肢の屈曲と伸展、手の開閉、口の開閉といった多様な基本的運動が誘発されます。
これを応用したある実験は、コンピューターをサルの脳幹に接続し、基本的運動を順次かつ並行に誘発するようにプログラムすることで、サルの体を意のままに操作できることを示しました。
驚くべきことに、この実験は50年以上も前の1960年代に実施されました。
移動運動と生体ロボット
移動運動
歩行や遊泳などの移動運動は、ほとんどの動物と人間に共通する基本的な運動行動で、その大部分が脊髄の実行回路の活動によって達成されます。(Leiras et al. 2022)
この脊髄回路に対する直接的な中枢制御は脳幹にあり、脳幹は移動運動の開始、速度、終了、方向を制御しています。
脳幹の電気刺激で動物に歩行、走行、遊泳、旋回などの移動運動を誘発することができることが示されています。
生体ロボット
生物と人工物の融合はロボット工学における新たなトレンドで、融合したハイブリッドな動物は生体ロボット、動物サイボーグ、または動物ロボットと呼ばれています。(Romano et al. 2018)
生体ロボットを実現する一つのアプローチは、脳電気刺激を与えて動物の移動運動を制御することです。
これにより、動物をラジコンカーのように無線で遠隔操作したり、動物をコンピュータと接続して指定経路を自律的に走行させることができるようになります。
自律神経系
自律神経系の運動部門は、呼吸、心臓、血管運動、消化、内分泌など、さまざまな体内の生理機能を制御します。(Beissner et al. 2013)
自律神経機能の最も重要な制御中枢のいくつかは脳幹に位置しています。
脳幹に加えて、視床下部、島皮質、帯状皮質、扁桃体が中枢の自律神経ネットワークの中核を形成しています。
自律神経系は心臓を制御しており、その直接的な入力は脳幹から生じます。(Silvani et al. 2016)
また脳幹には呼吸中枢が存在し、自動的かつ継続的な呼吸運動にはこの回路が必要不可欠です。(Saper et al. 2013, Bianchi and Gestreau 2009)
そして脳幹の電気刺激で、心停止、呼吸停止を引き起こすことができます。
また脳幹の電気刺激で嚥下、くしゃみ、咳、嘔吐、排尿、排便といった自律神経系の運動を誘発することができます。
身体操作
コンピュータによる運動制御
スタンフォード研究所 (現SRIインターナショナル) のローレンス・ピネオ博士は1966年、サルの脳幹をコンピューターに接続し、サルにプログラムされた一連の運動を行わせることに成功しました。(PINNEO 1966, Pinneo et al. 1972)
この実験は米海軍研究局から資金提供を受けており、内容から軍事的な応用を念頭に置いたものであると読み取れます。
この実験では、3匹のアカゲサルの脳幹・小脳をコンピューターにケーブルを介して接続し、コンピューターでこれらの運動神経を操作して、サルに複雑な運動を実行させることを試みました。
博士はまず、サルの食事、毛づくろい、移動、遊び、攻撃行動などの目標指向型の行動を分析し、それを構成要素である基本的運動に分解しました。
これらの基本的運動を順次かつ並行に誘発することで、協調した四肢の動きを生み出し、様々な複雑な運動をサルに行わせることができると、博士は考えました。
次に、さまざまな基本的運動を生み出すために脳幹と小脳のどの部位を刺激する必要があるのか、広範な脳機能マッピングが行われました。
電極は小脳核と脳幹の前庭神経核、網様体、その他多くの脳幹構造に配置され、電気刺激によって基本的運動を生み出す地点が200箇所以上見つかりました。
これらの基本的運動には、手首・肘・肩・足首・膝・股関節における四肢すべての屈曲と伸展、指の握り締めと広げ、微細な指の動き、口の開閉、舌の出し入れ、尾の巻き上げまたは横への動き、眼球の単独または同時の動き、瞳孔の散大、そして心拍数や呼吸数の変化といった多くの自律神経反応が含まれていました。

また、脳幹と小脳から得られる運動は、運動皮質の刺激によって引き起こされる運動よりもはるかに正確であることがわかりました。
その後、最大60個もの電極がサルの脳に慢性的に埋め込まれ、ケーブルを介してコンピューターに接続されました。

コンピューターは最大10個の電極を同時に操作することできました。
そして基本的運動の組み合わせをプログラムとして記述し、それをコンピューターに実行させました。
するとピネオ博士の思惑どおり、サルに様々な目標指向型の運動を行わせることができました。
これらには例えば、手足を伸ばして食べ物などの物体をつかんで素早く滑らかに口に運ぶ運動、木登りのように腕を外側と上側に伸ばす運動、尾の付け根を引っ掻くように腕を体の後ろに回して前後に動かす運動などがありました。
相同部位を刺激した時の3匹のサルの基本的運動は非常に類似しており、特定の一連の刺激により各サルに本質的に同じ一連の行動が生じました。また、異常に見える時間の流れ、すなわち「スローモーション」行動も得ることができました。
この一連の行動は望むだけ繰り返すことができ、観察者はその動きを、サルが自発的に同じ行動をとったときに生じた動きと区別することができませんでした。
プログラムと刺激の持続時間を変えるだけで、ほぼ無限の一連の運動のレパートリーを引き出せるようでした。
いずれの場合も、電気刺激はサルのあらゆる自発的な動きを無効にし、どんなサルの姿勢においても、またサルが眠っているか、起きているか、麻酔下にあるかに関係なく、本質的に同じ一連の運動を誘発することができました。

生体ロボット (移動運動の制御)
脳幹には電気刺激によって前進、左旋回、右旋回などの移動運動を誘発することができる部位が存在します。
これにより、動物をラジコンカーのように無線で遠隔操作したり、動物をコンピュータと接続して指定経路を自律的に走行させることができるようになります。
つまり、動物を生体ロボット化することができるようになります。
前進運動
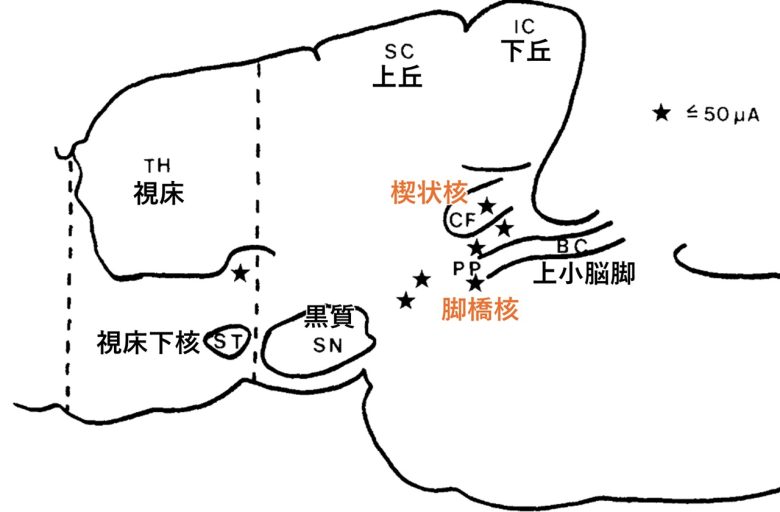
ロシアの研究者は、ネコの中脳の楔状核を電気刺激すると、歩行と走行を誘発できることを発見しました。(Shik et al. 1966, Fridman 1969)
刺激の強度が一定であれば、ネコは一定の速度で走り、非常に協調した動きをしました。
刺激の強さを調節することで走行速度を変えることができ、ネコは歩き方を歩きから走りへと変化させました。
これらの歩行と走行の基本的な特徴は、健常動物のものと似ていました。
サルを使った実験でも、楔状核の電気刺激で歩行と走行を誘発できることが示されています。(Eidelberg 1981)
カニクイザルの楔状核を電気刺激すると、歩行に似た、四肢のリズミカルな交互運動を誘発されました。
弱い刺激では、刺激開始から2~5秒後に反対側の後肢が動き始め、刺激終了後も2~3秒間動き続けました。
刺激を強くすると、通常まずもう片方の後肢が、次に前肢がゆっくりとした歩行に似たリズミカルな動きで動員されました。
さらに強い刺激を与えると、歩行速度が増加し、その後、全速走行に移行しました。
アーカンソー大学の研究者はラットを使った実験で、脳内で歩行を誘発する領域を広く探索しました。(Skinner and Garcia-Rill 1984)
すると楔状核の他、脚橋核からも歩行を誘発できることがわかりました。
この他、視床のすぐ下にも歩行を誘発できる部位がありました。
この実験でも刺激強度よって歩行速度を変えることができました。
弱い刺激では歩行、中程度の刺激では速歩、強い刺激では全速力の走りが誘発されました。
別の研究では、刺激周波数によっても歩行速度が変わることが示されています。(Caggiano et al. 2018)
刺激周波数を閾値周波数5 Hzから最適周波数50 Hzまで増加させると、移動速度が徐々に増加しました。
楔状核と脚橋核からなるこの領域は「中脳歩行誘発野」と呼ばれ、ヤツメウナギ、エイ、ネズミ、ブタ、サル、ヒトなど、さまざまな脊椎動物種で発見されています。(Noga and Whelan 2022)
旋回運動
日本の広島大学の研究では、金魚の内側縦束を電気刺激すると、前進運動だけでなく、旋回運動も誘発できることが示されました。(Kobayashi et al. 2009)
実験では、魚類用の極小の無線刺激装置が開発されました。これに浮きをつけ、金魚に装着しました。
金魚の内側縦束の正中線上の地点を電気刺激すると、前進運動が誘発されました。
一方、正中線から外れた部位を電気刺激すると、刺激側への旋回運動が誘発されました。
無線送信機から刺激装置にこれらの指示を送ることで、金魚をラジコンのおもちゃのように遠隔操作することができました。
中国の燕山大学の研究では、コイの小脳の電気刺激で左旋回、右旋回、前進、後退の4種類の移動運動を誘発できることが示されました。(Peng et al. 2011)
小脳の正中線上の上側の部位を刺激するとコイは前進し、下側の部位を刺激すると後退しました。
小脳の正中線の左側の部位を刺激するとコイは右に旋回し、右側の部位を刺激すると左に旋回しました。

ヤモリを使った実験では、中脳被蓋の広い領域から前進運動と旋回運動が誘発できることが示されました。(Wenbo et al. 2009)
中脳被蓋の正中線近くの部位の電気刺激は前進運動を誘発し、正中線から外れた部位の刺激は旋回運動を誘発しました。

ハトを使った実験では、脳電気刺激によって飛行運動を制御できることが示されています。(Yang et al. 2017)
視床の感覚中継核に電極が埋め込まれ、この核への電気刺激は左右の旋回コマンドとして機能しました。
方向指示はハトに装着した小型のコンピュータによって与えられました。
飛行中に一連の旋回コマンドが与えられると、ハトがそれに従って進路を変更することが確認されました。
生体ロボット
東京大学の研究者は動物の生体ロボット化の簡単な例として、触角刺激を使ってゴキブリを黒い線に沿って自律的に歩かせ続けることに成功しました。(Holzer and Shimoyama)
ゴキブリは触角刺激を受けると反対側に逃避します。
実験では、黒い線からはみ出すと触角への電気刺激を加える、小型の制御装置が開発されました。
これを装着したゴキブリは、黒い線からはみ出すたびに触角刺激を受け、線に戻るようになりました。
これにより自律的にゴキブリを黒い線に沿って歩かせることができました。
ニューヨーク州立大学ダウンステート医療センターは、ラットをラジコンカーのように無線で遠隔操作する技術を開発しました。(Talwar et al. 2002)
内側前脳束と、感覚皮質の左右のヒゲを表現する領域に、無線経由で電気刺激を加える電極が埋め込まれました。
ラットは内側前脳束の刺激を受けると前進し、感覚皮質の左または右のヒゲ領域への刺激を受け取るとその方向へ旋回しました。
これにより500m離れた地点からラットを遠隔操作することが可能になりました。
内側前脳束への刺激は、ラットを前進させるだけでなく、直面している段差を登り降りさせる動機付けにもなりました。
したがって、内側前脳束の刺激は、ラットを多種多様な複雑で新しく変化する地形を通過させるのに十分でした。
ラットはパイプや高架路、岩棚を容易に通り抜けることができ、木など十分な足掛かりのある面であれば、登ったり飛び降りたりするよう指示することができました。
また、ラットが崩壊した巨大なコンクリート瓦礫の山を組織的に探索するよう誘導したり、明るく照らされた開放的なアリーナなど、普段は避けるような環境をラットが通れるように誘導することもできました。
研究者はこのラットを「ロボラット」と名付け、この技術を動物の生体ロボット化に応用することへの熱意を表明しました。

ロボラットに触発された中国の浙江大学の研究グループは、ラットの脳をコンピュータに接続して、指定経路を自律的に走行させるシステムを開発しました。(Wang et al. 2015)
まずラットに小型カメラと、無線の脳刺激装置を取り付けました。
カメラはラットの前方の景色を撮影し、コンピュータにそれを無線で送信しました。
続いてコンピューターは標識を検出し、標識に応じて左折、右折、前進の3つのどれかの信号を無線で刺激装置に送信しました。
ニューヨーク州立大学の研究同様、前進は内側前脳束、左折と右折は感覚皮質のヒゲ領域への電気刺激で実現されました。
続いて樹木、道路、交差点を含む都市モデルを作成し、左右の矢印と人間の顔 (前進) の標識を設置しました。
ラットをこの模型に配置すると、実際にラットが設置された標識に従って都市モデル内を移動することが確認されました。

同研究グループはこれを進展させ、機械知能と生物知能の統合例として、迷路から脱出するラットをコンピューターで支援し、より早く迷路を解決できることを示しました。(Yu et al. 2016)
ラットは以下のようなルールでコンピュータからの支援、つまり脳電気刺激を受けました。
- 唯一の道への経路がある場合、コンピュータは最短経路を見つけ、左右のコマンドを送信して、ラットを唯一の道まで誘導。
- ラットが行き止まりに入ろうとしている場合、そのような動きを防ぐために左または右のコマンドを送信。
- ラットがループに陥っている場合、コンピュータは現在の目的地への最短経路を見つけ、左右のコマンドを送信して、ラットをその経路に沿って誘導。
ラットは14種類の迷路を解き、歩数、網羅率、所要時間が評価されました。
すると、コンピューターの支援を受けたロボラットは、迷路解決において通常のラットよりも優れた性能を示したことがわかりました。
ヘス博士の動物実験
スイスのチューリッヒ大学のヴァルター・ヘス博士は、ネコの間脳 (視床と視床下部) を電気刺激すると、様々な運動が誘発されることを発見しました。(Hess and Brügger 1943, Hess 1958)
頭部の運動
後交連から外側の領域を電気刺激すると、すぐにネコは頭を下げました。刺激が続くと、ネコは鼻を机にコツコツとぶつけました。
この領域を電気凝固すると、ネコは頭が持ち上がったままになってしまいました。
後視床下部の内側部を電気刺激すると、ネコは頭部を持ち上げ、場合によっては体の前部全体を持ち上げました。
この反応は刺激開始と同時に始まり、数回の急激な動きを経て、頭部を高く掲げた姿勢へと至りました。
ネコは横たわっているときに刺激を受けると、体を前に起こしました。刺激の強さが増すにつれてこの効果はさらに強まり、体の前部が床から持ち上げられ、ネコは後ろに倒れてしまいました。
ネコは倒れる際、背中から落ちないように、縦軸を中心にして素早く回転しました。

回転運動
回転反応は非常に頻繁に現れ、視床から視床下核にかけての領域から誘発されました。
より前部および背側の領域で誘発される回転運動は主に頭部に関連し、時にそれに限定されました。


しかし、より後方およびより腹側の領域を刺激すると、頭部だけでなく体の前部、最終的には全身に影響が及び、転がる運動へと容易く進展しました。
この効果は刺激が強いほど顕著になりました。
ネコが仰向けになるまで回転が進むと、ネコは人工的に誘導された動きの方向へぐいと身を素早く引き寄せ、即座に再び立ち上がりました。

四肢の運動
視床の腹側部を刺激すると、引き上げ、内転、外転という、3種類の前肢の運動がネコに生じました。
刺激周波数に合わせて前肢が上下に動く現象も観察されました。

顔面運動
顔面における運動反応は非常に頻繁に観察され、耳の筋肉、まぶた、ヒゲでは簡単に得られました。
圧倒的に最も一般的な反応は、反対側のヒゲが後向きに反り返ることでした。
刺激効果が強い場合、例えば、ヒゲから上唇まで動きが広がり、上唇は反対側に引っ込みました。時には反対側の頬まで巻き込み、頬が持ち上がることもありました。
少し強い刺激によって同じ側のヒゲも動くことがよくありましたが、後向きではなく前向きでした。
耳にも同様の行動が観察され、最も一般的な反応は、反対側の耳を後ろに回し、同じ側の耳を前に回すことでした。
まぶたの動きは、ほとんど例外なく反対側で見られました。
時には同じ側まで動きが広がることもありましたが、その場合は常にかなり弱まりました。反対側のまぶたは完全に閉じているのに対し、同じ側のまぶたはわずかに閉じているだけで、刺激のリズムに合わせてわずかに瞬きしました。

顔面反応を誘発した部位は、間脳運動野の上位領域、すなわち視床放線が通過する領域に位置していました。
デルガドの動物実験
イェール大学の神経生理学者ホセ・デルガドは主にサルやネコ、一部では人間を対象に、脳電気刺激によって運動を誘発する実験を多数実施しました。(Delgado 1964, Delgado 1970)
脳のさまざまな部位を刺激すると、自発的な行動で観察される単純な動きのほとんどが誘発されました。
これには、顔をしかめる、目の開閉、口の開閉、舌を動かす、噛む、顔の収縮、耳を動かす、頭や体を回す・ねじる・曲げる・伸ばす、腕・脚・指を動かす、などが含まれました。
脳の領域と刺激パラメータに応じて、異常な反応、無秩序な動き、平衡感覚の喪失、てんかん様な痙攣も生じました。
例えば、サルの中脳蓋を電気刺激すると平衡感覚が完全に失われ、サルは崩れ落ち、縦軸を中心に時計回りに回転しました。

瞳孔の直径は、視床下部に接続された刺激装置の強度ノブを回すだけで、まるでカメラのように最大限の収縮から最大限の拡張まで意のままに調整できました。

脳表面の刺激
ネコの右側の運動皮質に電気刺激を与えると、左側の後肢が屈曲し、その運動の大きさは適用された強度に比例しました。
例えば、ネコが四つ足で立っているときに、1.2 mAの強度では後ろ肢が地面からわずかに浮く屈曲が生じました。1.5 mAでは肢が約4 cm上がり、1.8 mAで肢の屈曲が完遂しました。

サルでも運動皮質の電気刺激により、ネコと同様、刺激とは反対側に運動が誘発されました。
人間の被験者では、左頭頂皮質を電気刺激すると、最初の2本の指の収縮から始まり、他の指の屈曲が続く、右手の屈曲が誘発されました。
その後、5秒間の刺激の残りの間、握りこぶしは維持されました。
この効果は不快でも不安でもなく、進行中の行動や自発的な会話を妨げることなく発現しました。
被験者に刺激が来ることを警告し、指を伸ばしたままにするよう指示したところ、彼は誘発された動きを阻止できず、「先生、あなたの電気は私の意志よりも強いようですね」と発言しました。
被験者が自発的に手を使って、たとえば雑誌のページをめくっているときにこの刺激が加えられると、この動作は妨げられませんでしたが、誘発された手の屈曲により自発的な動作が歪められ、ページがくしゃくしゃになり破れてしまいました。
運動皮質の電気刺激は正確で巧みなな動きを誘発せず、すべてのケースで誘発された反応は不器用で異常なものでした。
脳深部の刺激
大脳辺縁系や視床など、脳深部の領域の刺激によって引き起こされる運動は、通常、運動皮質の刺激によって生じるものよりも複雑で、目的があるようにみえました。
たとえば、人間の被験者では、内包の吻側部を電気刺激すると、被験者が何かを探しているかのように、頭を回転させ、体をいずれかの側にゆっくりと移動するという、よく方向づけられた、明らかに正常な一連の動作が見られました。
この刺激は2日間にわたって6回繰り返され、同様の結果が得られました。
被験者は誘発された活動は自発的なものであるとみなし、常にそれについて合理的な説明をしました。
「何をしているのですか?」と尋ねると、「スリッパを探している」「音が聞こえた」「落ち着かない」「ベッドの下を探している」といった答えが返ってきました。
刺激の再現性
脳のある地点に刺激を与えると、数時間、数か月、あるいは数年の間隔を置いて、同様の効果が一貫して誘発されました。(Delgado 1959)
再現可能な効果には、頭を回すなどの身体的反応、瞳孔の収縮や拡張などの自律神経反応、恐怖、逃走、発声などの行動的な効果などがありました。
サルの左嗅脳溝の電気刺激により、頭が右を向き、歯を見せずに口が開き、左手で顔の左側を掻く動作が誘発されました。
1時間の間に同じ地点を60回刺激したところ、1分ごとに同じ反応が繰り返されました。

刺激の開始時にサルが食事中または歩行中だった場合、その行動は中断され、口を開ける・頭を回す・掻くという効果が現れ、その後、刺激が止まるとすぐにサルは元の行動を再開しました。
左側の黒質と動眼神経核の間の地点を電気刺激すると、左眼輪筋の収縮と左眼の右への回旋が誘発されました。
4年2か月の間にわたり、この地点は100回以上刺激されましたが、誘発効果は一定でした。
右被殻の内包前脚に近い点を刺激すると、左脚の伸展、左手と左腕の屈曲、頭部と胴体の左への回旋と若干の屈曲、両眼の左への回旋が誘発されました。
この動作も長期間に渡って再現が可能でした。
走りの誘発
サルの海馬采への電気刺激により即座に荒ぶる走り反応が誘発され、この反応は刺激が加えられるたびに例外なく現れました。(Delgado 1967)
動物は刺激時間中ずっと、ケージ内を猛スピードで左右に走り回り、ケージの側面を半分ほど登っては勢いよく跳ね返りました。
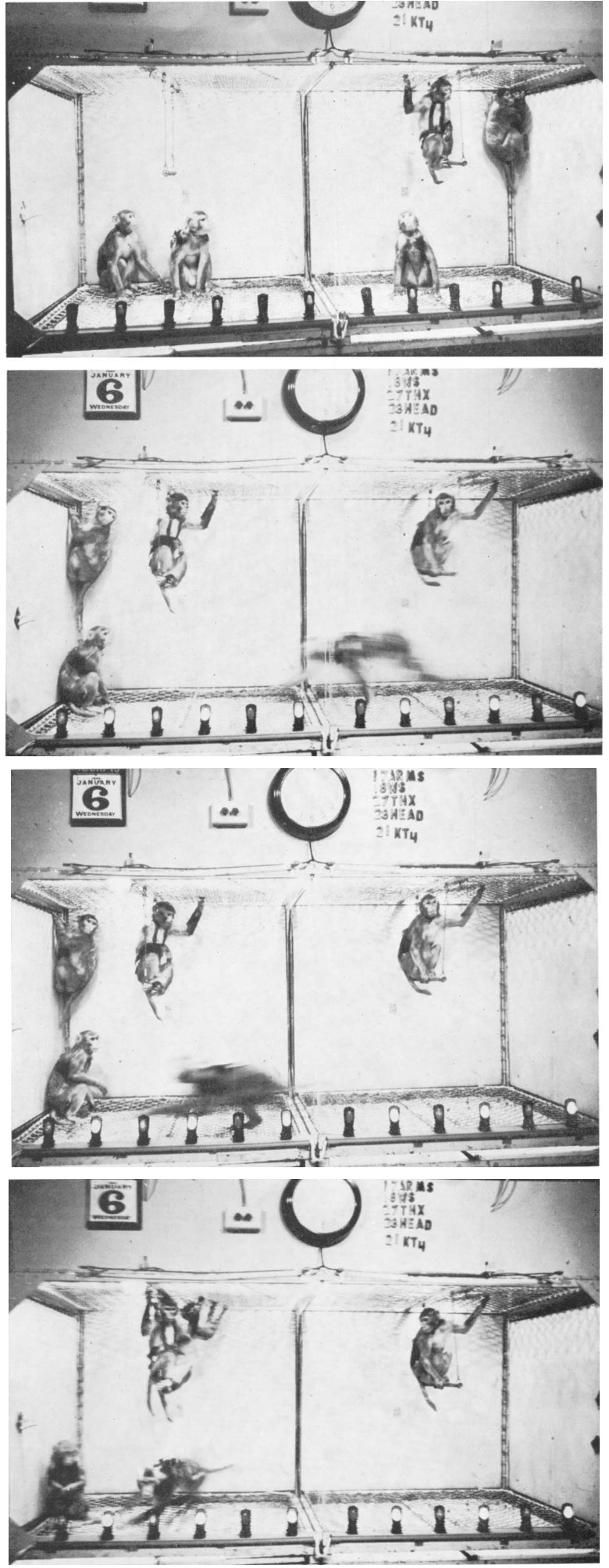
動きの協調性は優れており、方向性はよく保たれていました。
動物は他のコロニーの動物との衝突を避けることができ、時折1匹が轢かれても、争いや攻撃は起こりませんでした。
数回の試行の後、ケージの床全体が走っているサルに譲られ、他のサルはブランコに座ったり、隅にぶら下がったりしていました。
刺激が終わるとすぐに、刺激を受けたサルは座り込みました。
刺激中に餌を与えても、サルはそれに全く注意を払わず、走り回り続けました。脅かしすら効果がありませんでした。
デルガドは、ケージ内のレバーが押されると、実験サルの海馬采に電気刺激が与えられるようにしました。(Delgado 1970)
すると1匹のサルがこのレバーを押すことを学習し、実験動物への脳電気刺激が開始されました。
この刺激が繰り返されるうちに、操作サルのレバー押しに対して、実験サルが条件付けられました。
つまり、実験用のサルは、操作サルがレバーに近づくとすぐに落ち着きを失い、走り出す準備をしてケージの隅に立つようになったのです。

呼吸停止・無呼吸
呼吸停止
脳幹の電気刺激で永久的な呼吸停止を引き起こすことができます。
脳幹
フランスのポール・セザンヌ大学 (現エクス=マルセイユ大学) の研究では、ネコの脳幹を電気刺激すると、永久的な呼吸停止が生じることが発見されました。(Bassal and Bianchi 1982)
実験では呼吸に影響を与えるネコの様々な脳領域が探索されました。
横隔膜神経の活動開始は、一次感覚皮質、水道周囲灰白質、網様体、腕傍核、前庭神経核、小脳室頂核の電気刺激への電気刺激によって得られました。
特に水道周囲灰白質や網様体などの脳幹構造では、刺激が1~5 Hzの周波数で繰り返されると、呼吸が刺激周波数に同調しました。


横隔膜神経の活動停止は、運動皮質と内臓皮質、内包、錐体路、赤核、赤核脊髄路、三叉神経の運動核への電気刺激によって得られました。
特に錐体路、赤核脊髄路などの脳幹構造では、刺激が1~10 Hz の周波数で繰り返されると、呼吸が弱まるか、あるいは永久的に停止しました。


無呼吸
その他の脳領域からも、永久的な呼吸停止までいかずとも一時的な呼吸停止、つまり無呼吸を誘発できることが示されています。
眼窩前頭皮質
ボストンのマサチューセッツ総合病院のウィリアム・スウィートは、ネコとサルの眼窩前頭皮質の電気刺激で無呼吸が生じることを発見しました。(Bailey and Sweet 1940)
弱い電気刺激では呼吸が抑制され、強い刺激では呼吸が停止しました。
呼吸停止では、刺激開始から1秒未満で呼気が5~10秒間停止し、ゆっくりとした浅い呼吸で離脱しました。
そして刺激終了後には大きな吸気が発生し、正常な呼吸数と振幅へ即時に回復しました。
ネコとサルで基本的に反応は同じでした。
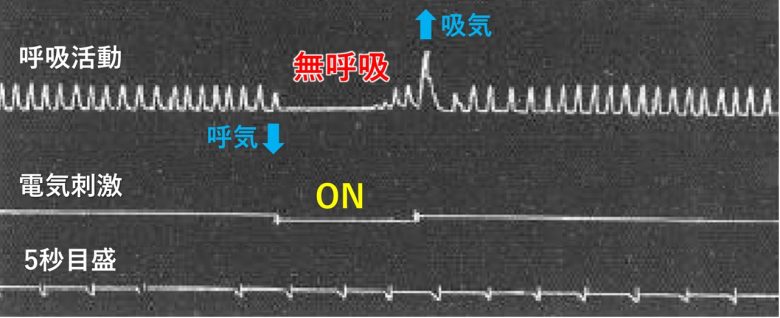
ボストン精神病院では、ロボトミー手術を控えた精神病患者に電気刺激を与えて無呼吸を誘発する人体実験が行われました。(CHAPMAN 1949)
患者7名は統合失調症、3名は混合性精神病でした。
眼窩前頭皮質への電気刺激により、7人の患者の呼吸が抑制または停止しました。
呼吸への影響は電気刺激から1~2秒以内に発生しました。
呼吸の回復は通常、刺激終了から2~4秒後に起こりましたが、2名の患者では3回の刺激後、それぞれ40秒間も無呼吸が続きました。
これらの反応は同じ部位の刺激によって再現することができました。
前帯状皮質
前帯状皮質の電気刺激でも無呼吸が生じることが示されています。(Smith 1945)
アカゲザルの前帯状皮質に電気刺激を加えると、わずかな遅延の後、呼吸が停止しました。
呼吸運動は刺激の持続時間が数秒を超えない限り、刺激期間中ずっと停止されました。
停止は呼気において生じ、刺激終了後も1~2秒続くことがありました。
その後呼吸が再び始まり、最初は浅いものの徐々に深くなり、数秒以内に刺激前のレベルに到達しました。
多くの場合、無呼吸に続いて深呼吸が起こり、その後呼吸はすぐに正常に戻りました。
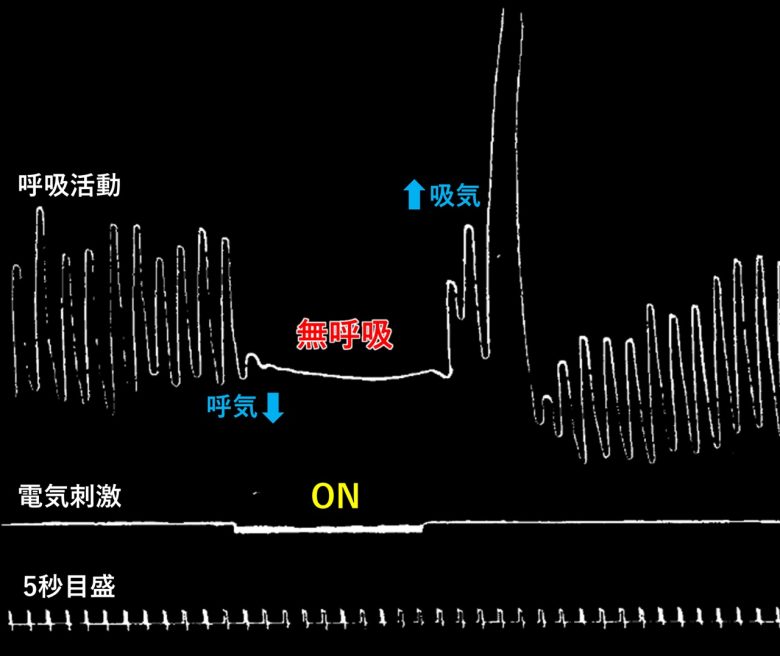
コロンビア大学では、前帯状皮質の電気刺激が自律神経系に与える影響を調べる人体実験が行われました。(Pool and Ransohoff 1949)
被験者はロボトミー手術を控えた11名の統合失調症の患者でした。
前帯状皮質に電気刺激を加えると、2名に無呼吸が生じました。
まず呼吸数が低下し、刺激の終了後、約30秒間の完全な無呼吸が続きました。
扁桃体
オスロ大学のビルガー・カーダ博士は、扁桃体の電気刺激でも無呼吸が生じることを示しました。(Kaada et al. 1954)
ネコの扁桃体の前内側部に電気刺激を与えると、呼吸運動が即座に抑制されました。無呼吸の間、胸郭は通常、呼気の位置をとりました。
呼吸運動は連続的に刺激しても通常は40~60秒以上停止させることができず、その後は離脱が生じました。
しかし、刺激終了後、かなりの時間呼吸が停止する動物もおり、その場合は自発呼吸を促すために人工呼吸が必要でした。
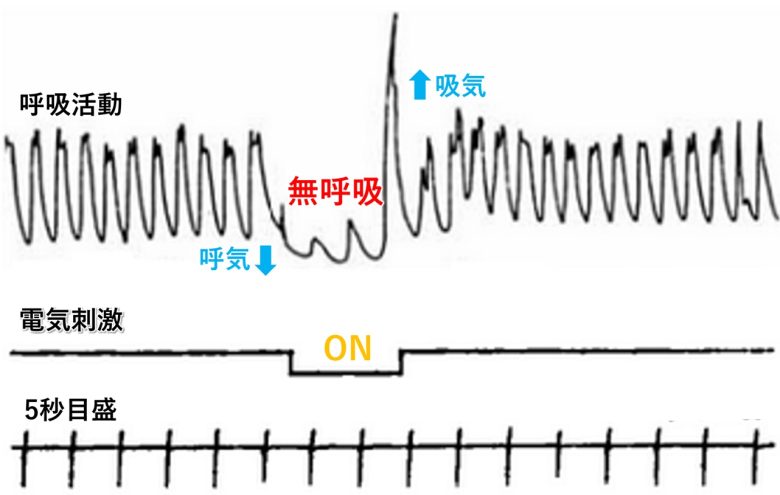
その他の領域
カーダ博士は続けて、サルの大脳皮質で呼吸反応を引き起こす領域を広く探索しました。(Kaada et al. 1949)
すると大脳皮質の幅広い領域への電気刺激で、呼吸の抑制あるいは停止を誘発できることがわかりました。
これらの反応を誘発した部位は、側頭極、島皮質の前部、眼窩前頭皮質の後部、前帯状皮質、および中隔野に渡って連続的に広がっていました。
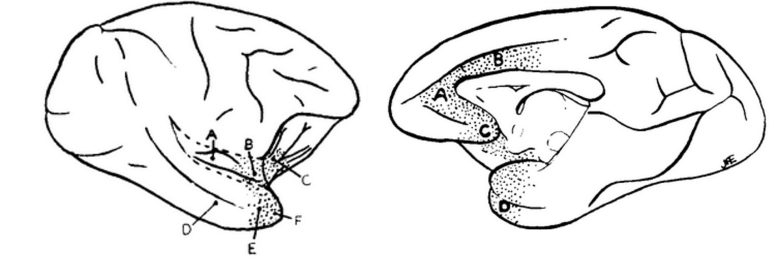
左:脳の内側領域、右:脳の外側領域。
弱い電気刺激では呼吸が抑制され、強い電気刺激では呼吸が停止しました。
完全な無呼吸が起こるとき、胸郭は呼気の位置をとりました。
無呼吸は25~30秒以上持続することはなく、その後は刺激を継続しても呼吸運動が再確立されました。
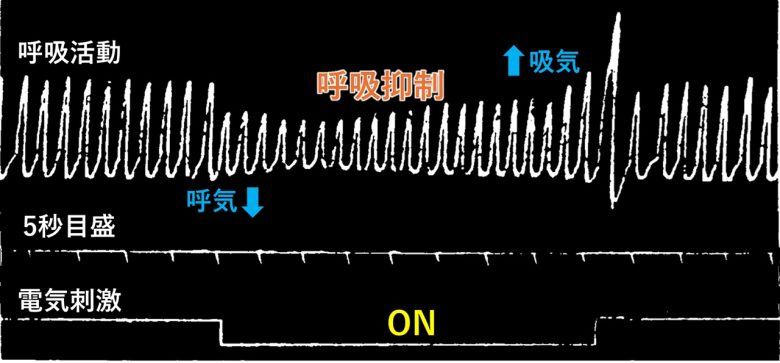

心停止・不整脈
心停止
脳幹の電気刺激で心停止を引き起こすことができます。
また、島皮質への複雑なパターンの電気刺激によって心停止が生じることが確認されています。
脳幹
マギル大学の研究では、ラットの脳幹の電気刺激で心停止が生じることが発見されました。(Malmo and Mundl 1983)
実験は人間で鎮痛作用が確認されていた、中脳領域の電気刺激の副作用を調査する目的で行われました。
するとラットの中脳の上小脳脚を電気刺激すると、ラットの心臓が停止することがわかりました。
これは同じ地点の電気刺激により再現できました。
酸素が直ちに鼻腔内に投与され、その後の回復は平均動脈圧の急上昇によって特徴づけられました。
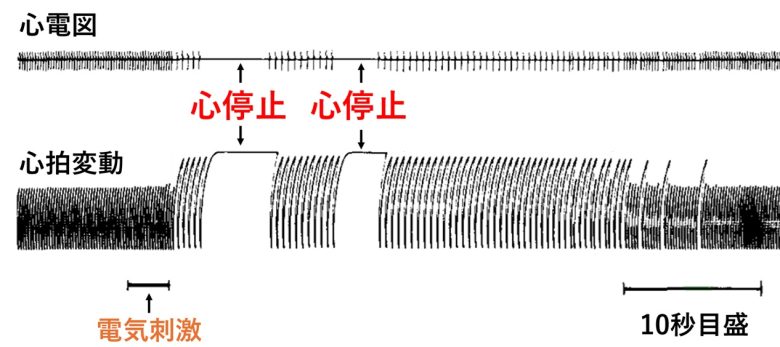
その他の中脳部位の電気刺激では、血圧の増減や呼吸の変化が起きる事が示されました。
島皮質
不整脈や心停止に大脳皮質が関与していることが示唆されています。(Oppenheimer and Cechetto 1990)
ジョン・ホプキンズ大学のスティーブン・オッペンハイマー博士は、自律神経の制御に関与している島皮質に目をつけ、この領域の電気刺激の実験を行いました。
するとラットの心拍に同期させて島皮質を電気刺激すると、頻脈と徐脈が生じることがわかりました。
頻脈は島皮質後部の吻側部から誘発され、刺激から平均8秒で始まり、平均15拍/分の心拍数上昇を引き起こしました。
徐脈は島皮質後部の尾側部から誘発され、刺激から平均9秒で始まり、平均16拍/分の心拍数低下を引き起こしました。
刺激は心拍数のみに影響を与え、他のパラメータには影響を与えませんでした。
博士は研究を推し進め、島皮質の電気刺激で不整脈からの心停止が可能かどうか調査しました。(Oppenheimer et al. 1991)
ラットの心拍に同期して、5ミリ秒の3つのパルスが心電図のT波の100ミリ秒前に、2周期毎に1回、島皮質の頻脈を引き起こす領域に送られました。
これにより心臓の電気的不安定性が最大になる期間中に交感神経活動のバーストが生じると推定されました。
すると刺激を受けたラット7匹すべてに、8時間以内に顕著な心電図の変化と不整脈が発生しました。
数時間にわたって心電図異常が徐々に進行し、続いて心室性の期外収縮が現れ、完全な心ブロックが発生しました。
心拍リズムはますます不規則になり、ついには顕著な徐脈が観察されました。
最終的に心停止が生じ、ラットは死亡しました。
不整脈
その他の脳領域からも、心停止までいかずとも期外収縮などの不整脈を誘発できることが示されています。
視床下部
アムステルダム大学の研究では、ネコの視床下部の電気刺激で不整脈が起きる事が発見されました。(Korteweg et al. 1957)
視床下部の前部に電気刺激を加えると徐脈が発生し、心拍が190拍/分から100拍/分まで即座に減少しました。
視床下部の後部に電気刺激を加えると高血圧が生じ、約125 mmHgから200 mmHgへの顕著な上昇がみられました。また刺激終了後に、期外収縮が頻繁に発生しました。
カリフォルニア大学の研究でも、ネコの視床下部の電気刺激で不整脈が起きる事が示されました。(Weinberg and Fuster 1960)
主に視床下部の後部および外側領域の電気刺激により、不整脈の多様な症状が得られました。
これらには、房室解離、期外収縮、頻脈、およびWPW症候群などがありました。
マギル大学の研究でも、ネコの視床下部の電気刺激で不整脈が起きる事が示されました。(Melville et al. 1963)
視床下部の前部の刺激では徐脈、外側視床下部の刺激では頻脈が生じました。
また双方のケースで期外収縮が観察されました。
前帯状皮質
コロンビア大学では、前帯状皮質の電気刺激が人間の自律神経系に与える影響を調べる人体実験が行われました。(Pool and Ransohoff 1949)
被験者はロボトミー手術を控えた11名の統合失調症の患者でした。
帯状皮質の電気刺激により、徐脈と頻脈が生じました。
徐脈は7人にみられ、最大で脈拍が40回/分低下しました。
4人の患者では、これらの変化は刺激期間を超えて持続しました。この効果の最長時間は10分間で、この症例では脈拍が著しく不規則になり、期外収縮が発生しました。
一方、頻脈は3人にみられ、最大で脈拍が40回/分上昇しました。
また血圧の上昇が8人に生じ、最大で収縮期血圧の60 mmHgの上昇と拡張期血圧の30 mmHgの上昇がみられました。
これらの変化は通常、刺激が加えられている間のみ持続しました。
その他の脳領域
ノルウェーの神経生理学者カール・セム=ヤコブセンも統合失調症やパーキンソン病の患者を使った人体実験で、脳電気刺激により不整脈などの心血管の変化が起きる事を示しました。(Sem-Jacobsen 1968)
前頭葉および脳の中心部を電気刺激すると、心拍数の増減、血圧の増減、期外収縮などの心血管変化が生じました。
また顔面紅潮あるいは顔面蒼白も生じ、時にはより複雑な反応が得られることもありました。
以下はある患者で生じた不整脈の描写です。
「彼の胸と体に流れ込んでいます。彼はとても気に入っています。彼は汗をかき、脈拍数が42に下がると青ざめ、ひどい吐き気が束の間生じますが、彼は極めて心地よい感覚を味わっています。幸福感が支配的ですが、吐き気の感覚がスパイスのようにそれに加わります。」
生理現象
脳幹の電気刺激によって嚥下、くしゃみ、咳、嘔吐、排尿、排便などの生理現象を引き起こすことができます。
その他、視床下部や扁桃体などの大脳辺縁系からも同様の現象が起きる事が観察されています。
脳幹
嚥下
ネコの脳幹尾部への電気刺激は、嚥下を誘発しました。(Miller 1972)
嚥下を誘発する電極部位のほとんどは、孤束核の腹側および外側に位置する背側延髄網様体に集中していました。
完全な嚥下パターンは、10~30 Hzの最適周波数で誘発されました。
刺激強度を増加させると、刺激中に筋肉に「テタニー様の収縮」が起こり、その後、数回の嚥下が続きました。
くしゃみ
ネコの脳幹下部への電気刺激は再現性よく、くしゃみを誘発しました。(Nonaka et al. 1990)
くしゃみ誘発部位は、脊髄三叉神経核の腹内側部および隣接する橋延髄の外側網様体に沿って位置していました。
1~50 Hzの複数の刺激周波数が試され、10 Hzが最も効果的であることがわかりました。
咳
ネコの延髄の孤束核の電気刺激は、咳を誘発しました。(MORI and SAKAI 1972)
脳幹誘発性の咳は、末梢神経の刺激で誘発されたものと明確な違いはみられませんでした。
どちらの反応も、1秒間に1~2回の割合で生じる、一連の痙攣性の吸気と高振幅の爆発的な呼気から構成され、これは以下の呼吸波形に見られました。
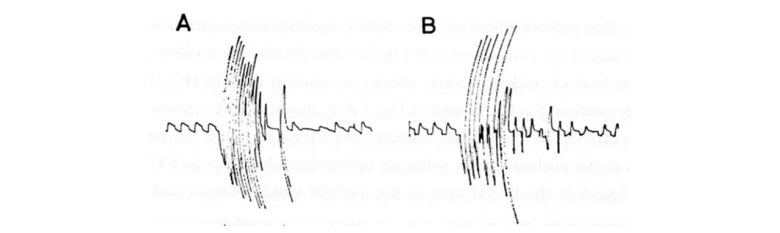
嘔吐
ネコの延髄の特定領域への電気刺激は、嘔吐を誘発しました。(WANG 1950)
この領域は、孤束とその核を含む外側網様体の背外側縁に相当しました。
最適な刺激周波数は50 Hz付近でした。
イヌにおいてこの脳領域を破壊すると、嘔吐誘発物質を与えても嘔吐しなくなりました。
排尿
ネコの橋の特定領域への電気刺激は、排尿を誘発しました。(Holstege et al. 1986)
電極部位は、橋の背外側被蓋の内側部に位置していました。
この領域への刺激は、骨盤底筋電図と尿道内圧の即時的な低下を引き起こし、2秒の遅延後に膀胱内圧の上昇が起こり、正常な排尿を模倣しました。
しかし、刺激による完全な排尿はたまにしか起きませんでした。
排便
イヌの橋上部の電気刺激は、排便を誘発しました。(Okada et al. 1976, Fukuda et al. 1977)
排便促進に関係するニューロンは、橋上部の三叉神経運動核の内側に位置する外側網様体に局在していました。
対照的に、この領域の電気凝固により、排便促進の活動が著明に減弱あるいは消失しました。
視床と視床下部
ヘス博士はネコの間脳 (視床と視床下部) の様々な領域に電気刺激を加えると、様々な生理現象が生じる事を発見しました。(Hess 1958)
あえぎ
頻呼吸は前視床下部の中間部と外側部から得られました。
パンティング (ハアハアというあえぎ) は、はるかに広い領域から誘発され、中隔、視索前野、外側および後視床下部に散在していました。
よだれ
よだれを誘発する領域は、あえぎを誘発する領域とほぼ同じ領域でした。
この一致には例外があり、視床の腹側部および視索前野の内側部では、あえぎのみが誘発されました。
一方、腹側視床の前縁、特に外側領域の地点を刺激すると、よだれのみが観察されました。
くしゃみ
くしゃみを誘発する部位は、中隔、特に隣接する脳室との境界部にかなりまばらに分布していました。
くしゃみは視索前野からも誘発されることがありました。
嘔吐
嘔吐を引き起こす刺激点はそれほど多くなく、視床下部、すなわち視索上野にあり、つまり視索前野の「くしゃみ点」に直接隣接しており、また乳頭体周辺の領域にもありました。
視床の腹側核にも嘔吐が誘発される刺激点が2つありました。
視床下部では、胃から大量の嘔吐物が排出される本格的な嘔吐が起こりました。
しかし視床では、反応は嘔吐運動の特徴を備えていましたが、内容物は吐瀉されませんでした。


舐め
舐める運動が誘発された部位は、視床腹側核の内側部にありました。
腹側核への刺激によって引き起こされた舐める運動は、ネコが何かを舐め上げたり、舐めたりしているように見えました。
頭を下げながら舌は規則的なリズムで動くことが多かったですが、舌が空を切る場合もありました。
隣接する部位の刺激によって誘発された舐める運動は、まっすぐに伸びたり、斜め上方に伸びたりするなど、異なる形で現れました。
匂い嗅ぎ
匂い嗅ぎは、外側視索前野と外側視床下部を含む領域の刺激によって得られました。
特に、内側前脳束に沿った領域は、最も低い刺激強度で匂い嗅ぎを生じました。

排尿と排便
排尿と排便は、地形的に区別できない同じ領域の電気刺激で誘発されました。
この領域は明確に限定されており、中隔野から始まり、その後縁が最も反応しやすいようで、視索前野と視索上野の背側部分を横切り、前視床下部へと進みました。
扁桃体
カーダ博士は、ネコの扁桃体への電気刺激でも、いくつかの生理現象が起きる事を示しました。(Kaada et al. 1954)
舐める、嗅ぐ、噛む
舐める、嗅ぐ、噛むという動作は、扁桃体の内側部と前部から最も頻繁に得られました。
ネコは床に沿って舐めたり嗅いだりするか、反対側を探すようにゆっくりと頭を動かし、自分自身を舐めたり嗅いだりしていました。
舐めと嗅ぎは刺激開始後の数秒以内に始まり、刺激が続く限り続きましたが、噛みは通常10~15秒遅れて発生しました。
排尿と排便
排尿は扁桃体の前内側核群と基底核から得られました。
排尿は通常、刺激の開始後の数秒以内に起こり、同じ地点を刺激することで繰り返すことができました。
排便は扁桃体の前部のある一点を刺激した場合にのみ起こりました。
毛の逆立ち
扁桃体の基底核の内側部の地点を刺激すると、2匹のネコで毛の逆立ちのが観察されました。

よだれ
扁桃体の前内側部からはよだれが得られました。
刺激を受けたネコは、粘液性の濃い唾液を大量に分泌しました。唾液の分泌は、多くの場合、舐める、嚥下、または噛むといった動作と関連していました。
その他の脳領域
カール・セム=ヤコブセンは統合失調症やパーキンソン病の患者を使った人体実験で、脳電気刺激により様々な生理現象が起きる事を示しました。(Sem-Jacobsen 1968)
生じた反応は、呼吸の変化、深呼吸、咳、発汗、吐き気、嘔吐、涙目、尿意など、多岐に渡りました。
反応の大部分は脳の前部にある電極から得られました。
呼吸の変化は刺激中に観察されましたが、深呼吸は刺激の終わりに最も頻繁に観察されました。(反応の有り様からして、実際には無呼吸を引き起こしたことが強く示唆されます。)
吐き気があると申告した患者は、その後嘔吐することもありました。吐き気は刺激終了後も続くことがよくありました。
尿意は「トイレに行かないと」「お願い、すぐに」などと表現されました。
しかし、刺激をやめると、患者は尿意を感じなくなり、「急がなくていい」「続けていいよ、急いでないから」などと自発的に発言しました。
