抑制
尾状核の電気刺激で、攻撃性抑制、運動停止、睡眠、無力症 (※)、拒食、記憶喪失など、さまざまな抑制効果を引き起こすことができます。
筋緊張の喪失を伴う不活性な状態。
中隔野の電気刺激でも攻撃性が抑制され、前障では意識障害が生じることが示されています。
これを悪用すると、不正に怒りや不満を募らせる人々を抑制し、集会やデモ、抗議活動を未然に封じることも可能になると思われます。
少なくとも実際に、チンパンジーの脳とコンピュータを無線で接続し、脳波から興奮状態が検出されると脳を遠隔から刺激しておとなしくさせるという実験が、50年以上も前にすでに行われていました。
目次All_Pages
回路
尾状核
大脳基底核は随意運動の制御に必須の神経回路で、線条体はその入力部に相当します。(Báez-Mendoza and Schultz 2013)
線条体は尾状核、被殻、腹側線条体・側坐核の3つの核から構成されています。
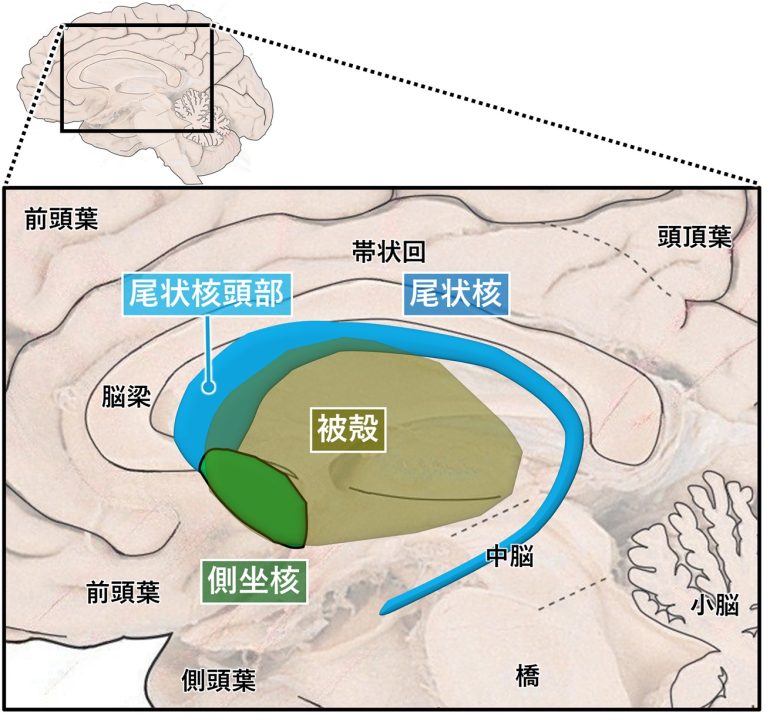
線条体の構成
(脳画像はVanderah 2018から)
尾状核の機能的役割は、一般的に大脳基底核の神経回路の構成要素として説明されることが多いですが、独自機能をもつ独立した核という観点から説明する研究者もいます。(Villablanca 2011)
尾状核を損傷すると、人間や動物が衝動的になり、強迫行動を示し、抑制が効かなくなることが知られています。
尾状核を両方除去されたネコは人間、他のネコ、または任意の物体に執拗に近づき、その後追いかけ、接触しようとしました。またネコはより覚醒し、睡眠不足の症状を示しました。
尾状核を両方除去されたサルは活動過多になり、ケージ内を強迫的に前後に行き来しました。数週間後、この歩き方はケージ内をぐるぐる回る行動に変化しました。
両側の尾状核頭部を損傷した女性には、以前の状態とは相容れない異常行動がみられるようになりました。(Richfield et al. 1987)
これらはほとんどが脱抑制の症状で、下品さ、衝動性、苛立ち、暴力的な爆発、多飲症と夜尿症、徘徊、食欲増進、性欲過剰、万引きや露出といった軽犯罪、不衛生などがありました。
注意欠陥多動性障害 (ADHD) の児童の尾状核頭部を正常児と比較したところ、左側が有意に小さいことがわかりました。(Hynd et al. 1993)
逆に尾状核の電気刺激では、損傷による脱抑制とは真逆の効果、つまり抑制が生じます。
尾状核、特に尾状核頭部の電気刺激すると、攻撃性や食欲を抑制し、実行中の運動を停止し、睡眠や無力症を誘発することが示されています。
中隔野、前障
中隔野に腫瘍を有する患者は「中隔の激怒」と呼ばれる攻撃性の爆発と易怒性を示し、ネズミを用いた研究でこの観察結果は裏付けられています。(Menon et al. 2022)
逆に中隔野の電気刺激は攻撃性を抑制することが示されています。
前障は広範な大脳新皮質の領域の抑制性のニューロンを同期させ、睡眠や休息時に観察される脳波である徐波の生成に関与しています。(Narikiyo et al. 2020)
前障の電気刺激で人間に意識障害が起き、動物も似たような状態に陥ることが示されています。
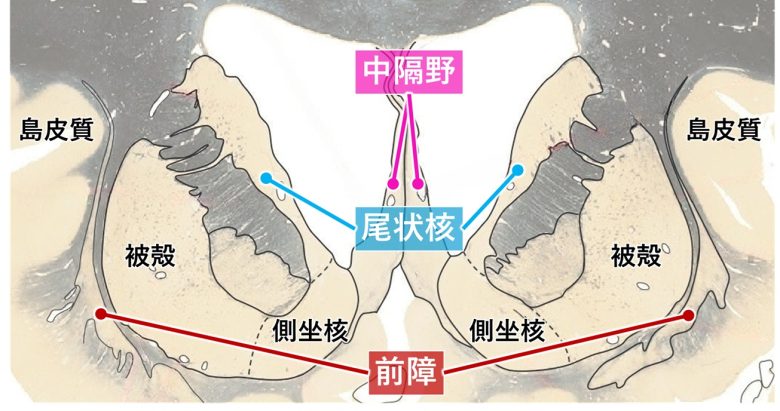
尾状核、中隔野、前障の位置
(脳画像はVanderah 2018から)
攻撃性抑制
尾状核や中隔野の電気刺激で、サルなどの動物の攻撃性が抑制されることが示されています。
デルガドの動物実験
イェール大学のスペイン人神経生理学者ホセ・デルガドは、スティモシーバーと名付けた無線脳刺激装置を発明しました。
彼はこの装置をアカゲザル、テナガザル、チンパンジー、雄牛に取り付け、攻撃性の抑制を目的とした多数の実験を手がけました。
雄牛
最も有名なのは、デルガド自身が雄牛と対峙し、遠隔操作によってその猛烈な突進を止めた実験です。(Marzullo 2017)
実験は1964年、スペインのコルドバにある農場で行われました。
雄牛の「ルセロ」にスティモシーバが取り付けられ、尾状核、視床、一次運動野を電波で遠隔刺激できるようにしました。
ルセロは手術後も力強い突進行動を続けました。
雄牛が闘牛士に突進しているときに、尾状核か視床を1 mAで電気刺激すると、雄牛はすぐに動きを止めました。
刺激を受けている間もルセロはじっとしていましたが、まばたきをし、規則的に呼吸していました。
電気刺激が止まると、雄牛は怒り狂って突進を再開しました。

続いてデルガドは無線送信機を片手に、自ら雄牛と対峙しました。
ルセロがデルガドに突進して2〜3メートル以内に近づくと、デルガドは刺激装置をオンにしました。
すると牛は激しく停止し、突然足が止まって大きな土埃が立ち、尾が下がり、頭が上がり、攻撃性が消えた状態を維持しました。
その後、デルガドは後ずさりし、リング内の木製の柵の後ろに身を隠し、刺激装置をオフにすると、牛はすぐに再び突進し、柵に衝突しました。

もう一匹の雄牛「カエタノ」の左尾状核の電気刺激では、左方向への回転運動が観察されました。
0.5 mAの刺激では雄牛は頭を左に動かし、0.7 mAでは完全に円を描いて回転しました。
右尾状核に刺激を与えると、同様の行動が観察されましたが、雄牛は反対方向、つまり右に回りました。
尾状核の刺激では全般的な抑制の他、回転運動が生じることが知られています。
チンパンジー
もう一つ有名なものは、チンパンジーの脳をコンピュータと無線接続し、興奮状態を自動検知して痛覚信号を送り、最終的に無気力化させた実験です。 (Delgado et al. 1970)
この実験の目的には「遠隔測定された脳波をコンピュータでパターン認識し、脳の特定領域に無線刺激を付随して誘発できるという原理を実証する」とあります。
明らかに群衆制御などへの応用を念頭においた軍事用途の実験であることが読み取れ、実際、この実験は米空軍の軍事施設内で行われました。
被験者は、「パディ」という名前の6歳の若い雄のチンパンジーでした。

スティモーシーバを取り付けた2匹のチンパンジー。左がパディで右がカルロス。(Delgado 1970)
パディの脳にスティモシーバが装着され、扁桃体と網様体に電極が埋め込まれました。
扁桃体の脳波は無線で送信され、研究所のコンピュータに解析され、紡錘波が検出されると自動で網様体への刺激信号が送られました。
扁桃体の紡錘波はチンパンジーの興奮と関連しており、網様体への電気刺激はパディに痛みを引き起こしました。
つまり、パディが興奮するとコンピューターが自動検知して痛覚信号が脳に送り込まれるというシステムが構築されました。

パディは米空軍の基地内に作られた人工島に、他のチンパンジーとももに放し飼いにされていました。
このシステムを運用開始すると、2時間でパディの扁桃体の紡錘波が半分に減少し、6日間でほぼ消失しました。
運用から10日後、視覚と嗅覚を使った識別能力の課題が実施されましたが、これはエラーなく実行されました。
しかし、重大な行動の変化が観察されました。
パディは非常に静かに座り、食べ物のペレットが運ばれてきても観察することなく、またしばしばそれを食べずに、与えられた課題を実行していました。
注意力と意欲の欠如は明らかで、パディの態度は実験前の通常の行動とは、非常に異なっていました。
彼は物静かになり、訪問者が訪ねてきたり、おいしい食べ物を持ってこられても興奮することがない、無気力な状態に陥ってしまいました。
また、デルガドはより直接的に、尾状核の電気刺激でチンパンジーの攻撃性を抑制できることを示しました。(Delgado 1970)
被験者のチンパンジーのカルロスはほとんどのチンパンジーと同様、かなり気難しく、罰をうけたり欲求不満になったり、あるいは単純に触られただけでも、簡単に癇癪を起していました。
彼は親しい人間から触られるのは好きでしたが、見知らぬ実験者から触られると、防御的で不安な反応を示していました。
彼は知人から触れられるのは好きでしたが、見知らぬ人からは嫌いでした。親しくない研究者が近づくと、防御的で不安な反応を示しました。
尾状核頭部へ電気刺激はこれらの防衛的な態度を抑制しました。
彼はなんの感情も示さず落ち着いて見え、何の騒々しい結果も起こさずに触ることができました。

アカゲザル
デルガドの脳電気刺激による攻撃性抑制の実験には、主にアカゲサルが用いられました。(Delgado 1964)
アカゲザルは獰猛な動物です。
手袋、棒、あるいは実験者の手が突然近づくと、サルはたいてい口を開けて歯を見せ、頭を前に動かし、特徴的な低声で鳴き、掴んだり引っかいたり噛んだりしようとしました。
尾状核のいくつの地点を電気刺激すると、サルは口を閉じて、このような攻撃的な態度を失いました。
サルに触れても安全で、指を口に入れても安全でした。顔に手袋や棒が近づくと、攻撃することなく手を使って押しのけました。
尾状核の電気刺激が止まるとすぐに、サルはいつもの凶暴な行動に戻りました。


デルガドは脳の電気刺激が、コロニー内のサルにどのような行動の変化をもたらすか実験しました。(Delgado 1963)
4匹のアカゲザルで構成されるコロニーにおいて、ボスのオスザルの「アリ」と、第二位のメスザルの「サラ」の脳の様々な部位に電極が設置されました。
水道周囲灰白質への電気刺激では、追いかけ、飛びつき、噛みつき、戦いの増加という、反社会的行動が双方に誘発されました。
逆に尾状核頭部への電気刺激は自発的な攻撃性を低下させました。アリは体格が大きく獰猛であったため、この効果はより鮮烈でした。
また、食物への関心の喪失や、飲水、ペレット摂取、歩き、拾い集め、毛づくろいといった様々な行動の抑制も引き起こしました。
尾状核刺激中はサルを素手で触ることができ、これによって何度もサルを捕まえることができました。
続いてボスザルのアリの尾状核頭部を電気刺激するレバーをコロニー内に設置し、下位のサルがどのような行動をとるか観察しました
するとアリと敵対関係にあった第三位のメスのサル「エルザ」が、威嚇された際にレバーを押してアリの攻撃性を低下させる行動が何度か観察されました。
レバーを押す間、彼女の注意はレバーではなくアリに向けられていました。これは珍しい行動でした。なぜなら、下位のサルが群れのボスをまっすぐに見つめると報復されるからです。

またデルガドは、中隔野の電気刺激によっても攻撃性を抑制できることを示しました。 (Rubinstein and Delgado 1963)
拘束されたサルを革手袋で脅すと、実験者に対して攻撃的な行動が引き起こされました。
サルは歯をむき出しにし、手袋を掴んで引っ張ろうとし、大きな声で鳴き、可能ならば激しく手袋を噛みました。
中隔野を電気刺激すると、これらの攻撃的な反応が抑制されました。
サルは近づいてくる手袋をただ見つめるだけで、鳴いたり噛んだりといった興奮の兆候は見せませんでした。しかし、頭を片側に向け、手袋をゆっくりと押しのけようとする動きが見られました。
刺激を与えている間、サルの顔や口に素手で触れても安全でした。
テナガザル
デルガドはテナガザルを使った攻撃性抑制の実験も手掛けました。(Delgado 1975)
テナガザルを拘束椅子に座らせ、革手袋で顔を軽く撫でると、通常は歯を見せたり、手袋を掴んで引っ張ろうとしたり、鳴いたり、噛みついたりといった攻撃的な反応が引き起こされました。
しかし、尾状核の電気刺激中は、テナガザルは興奮や敵意の兆候を見せずに近づいてくる手袋を見つめ、口に触れても安全でした。

尾状核頭部の電気刺激は、脳の他の部位の電気刺激で発生した攻撃性を抑制することもできました。
椅子に拘束されたテナガザルにおいて、水道周囲灰白質への刺激は攻撃的な行動を引き起こしました。
これは、落ち着きのなさ、歯を見せること、掴んで噛もうとすること、そして鳴き声に表れました。
水道周囲灰白質への刺激の2秒前に尾状核の電気刺激が先行すると、刺激誘発性の攻撃的な行動は消失しました。
コロニーケージ内で自由にされたテナガザルでも同様の結果が得られました。
水道周囲灰白質に電気刺激を与えると、黒色のテナガザル「ゲイリー」は金色のテナガザル「コティ」に対して的確な攻撃を仕掛けました。
コティは、体格と力が劣るにもかかわらず、防御して反撃しました。通常、2秒間の短い刺激で10~60秒間継続する威嚇や戦闘が誘発されました。
しかし、尾状核への電気刺激によってゲイリーは即座に抑制され、戦いを止め、無邪気で平和な姿勢をとりました。


その他の動物実験
尾状核および中隔野の電気刺激による攻撃性の抑制は、複数の研究者によって裏付けられています。
ネコ
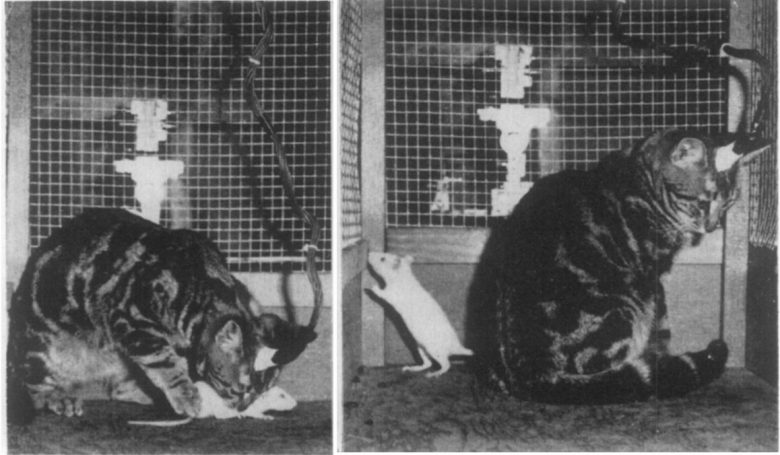
ダラウェア大学の研究者は、尾状核の電気刺激でネコの攻撃性を抑制しました。(Plumer and Siegel 1973)
ネコの視床下部への電気刺激は、55回のすべての試行 (100%) でラットへの攻撃行動を誘発しました。
尾状核の電気刺激がその20秒前に加えられると、53回の試行のうち7回 (11%) のみでラットへの攻撃行動が誘発されました。
また攻撃に要する時間も10~15秒長くなり、攻撃時間も非常に短くなりました。
ネコは尾状核に続いて視床下部の刺激を受けると、室内を周回し、ラットの追跡を開始しました。
しかし、ネコはラットの首に向かって凶暴な噛みつき攻撃を開始する代わりに、ラットを無視しました。攻撃しようとはせずにラットを踏みつけ乗り越えたりすることがよくありました。
ブリンマー大学の研究者は、中隔野の電気刺激でネコの攻撃性を抑制しました。(Thomas and Evans 1983)
まずネコの視床下部を刺激して、感情的な攻撃反応を誘発しました。
ネコは「シャー!」と鳴き、「カッ!」と怒声を発し、毛を逆立て、そして物体へ直接攻撃しました。
中隔野の電気刺激は、この感情的な行動に対して非常に強力な抑制効果を有していました。
ケージに中隔野を電気刺激を加える操作子 (ロッドあるいは台座) を設置すると、ネコはこれを作動させて不快な視床下部の効果を打ち消すことをすぐに学習しました。
ラット
ラットを使った実験では、中隔野の中でも外側中隔の電気刺激が攻撃性を抑制することが示されています。(Brayley and Albert 1977)
実験ではまず、視床下部に損傷を与えてラットを凶暴化させました。
次にラットの外側中隔、内側中隔、帯状皮質に電極を埋め込みました。
続いて刺激5分前、刺激中、および刺激5分後に、ラットの攻撃性を評価しました。
試験項目は、(a) 鼻先に鉛筆を差し出す、(b) 鉛筆で背中を強く叩く、(c) 鼻先に手を差し出す、(d) 棒で体の側面を優しく突く、(e) 尻尾をつまんで捕獲する、(f)体をつかむ、でした。
これらの試験に対する反応を0~3のスケールで定量化しました。
すると外側中隔への電気刺激でラットの攻撃性が約8割と劇的に低下しましたが、内側中隔と帯状皮質への刺激では目立った変化はみられませんでした。
攻撃性の低下
サル
サルを使った実験では、尾状核への電気刺激が痛みの感情的要素を軽減することが示されています。(Lineberry and Vierck 1975)
実験ではまずベニガオザルを拘束し、脚への電気ショックをうけたときに少なくとも2.25 kg以上の力でパネルを叩きつけ、電気ショックを止めるよう訓練されました。
そしてパネルに加えられた力が測定され、痛みの感情的な側面を反映する指標とみなされました。
脚の電気ショックはサルが止めない限り、15秒間続きました。
ベニガオザルの尾状核へ電気刺激は、脚の電気ショックから逃れるためにパネルに加えられた力を大幅に減少させました。
これは特に高強度の電気ショックにおいて顕著でした。
抑制効果は、尾状核刺激が脚の電気ショックに0~100ミリ秒先行した場合にのみ有効でした。
加えられた力
意識障害
前障の電気刺激で人間に意識障害が起き、動物も似たような状態に陥ることが示されています。
人間
ノルウェーの神経生理学者カール・セム=ヤコブセンは統合失調症やパーキンソン病の患者を対象に脳電気刺激の人体実験を繰り返していました。(Sem-Jacobsen 1968)
セム=ヤコブセンは脳の電気刺激によって、様々なレベルの意識の変化が生じることを発見しました。
反応を誘発した領域は「シルビウス溝と視床枕後部領域の間」と記述されており、かなり広範囲に渡っています。
この領域の中には前障も含まれます。
患者の意識の変化は、以下のカテゴリーに分類されました。
- 大発作を伴うまたは伴わない意識消失。
- 欠神 (短時間の意識喪失)
- 混乱している。咀嚼運動、唇鳴らし、嚥下。凝視。意識を失う寸前。
- 覚醒度の低下。正確な追従能力は5~25%の時間に制限される。
- 眠気。あくびの有無にかかわらず眠い。
- 意識がはっきりする。目覚める。完全に意識を取り戻し、思考を活性化する。
- 周囲のものが消えていく感覚。物が小さく見える。声が小さく聞こえる。失神に向かって漂っていく。
- 多重人格。多重人格の感覚。自分の外にいるような感覚。二人の人格に分裂している。
- 幻覚。視覚、聴覚、またはその他の形態の単純または複雑な幻覚で、他の精神病的行動は伴わない。
以下はある男性患者でみられた電気刺激による混乱の例です。
彼は第二次世界大戦中にカナダでノルウェー軍に所属し、英語を多少話しましたが、過去15年間は人里離れた山間の谷にある小さな農場で暮らしていました。
電気刺激中、記録のために患者は英語を使うよう依頼されました。
この患者は特定の電極を刺激された際に混乱状態になりました。彼は検査官の名前を言えず、英語で「あなたのお名前は覚えていません」と言いました。
また彼は事前に指示された通り、腕の反復運動を開始しましたが、なぜそうしているのか聞かれると、説明できませんでした。
彼は明らかに混乱していましたが、英語を話すことは覚えていました。
眠気は4人の患者に誘発されました。
これは「少し眠くなる。」 「より疲れを感じる。」 「くつろいで、眠くなる。」 などど説明されました。
特定の電極を刺激している間、患者の中には目を覚まさずに眠ってしまう人もいました。一方、他の電極を刺激すると、不快な副作用を引き起こすことなく、患者の睡眠状態が覚醒しました。
ワシントン大学のモハマド・コウベイシ博士は、前障の電気刺激で重度の意識障害が誘発された症例を報告しました。(Koubeissi et al. 2014)
患者はてんかんの54歳の女性で、病巣特定のために脳の電気刺激が行われていました。
左の前障と島皮質の間にある電極を刺激すると即座に意識障害が起こり、読み上げの停止、虚ろな眼差し、聴覚的・視覚的な指示への無反応、自発的な呼吸運動の鈍化などが伴いました。
これは10回の刺激中、10回再現されました。
刺激が止まるとすぐに患者は正常な状態に戻り、刺激期間中の出来事の記憶はありませんでした。
時折、誘発された意識障害は、1~2音節からなる、乏しく保続的で理解不能な言葉の発声を伴い、顔には混乱した表情がみられました。
ネコとネズミ
デューク大学の研究者は、前障の電気刺激でネコに不活性な状態を引き起こしました。(Gabor and Peele 1964)
刺激を受ける前、ネコはクリック音やケージを叩く音に即座に注意深く反応していました。
前障の電気刺激の開始から15~20秒後、動物は注意を引こうとする試みに反応しなくなりました。これは刺激期間を超えて続きました。
刺激が与えられるたびに、ネコは以下のような定型的な行動をとりました。
最初に、動物はしゃがんだ姿勢を取り、脚を体の下に曲げ、時折頭をわずかに反対に向けました。最後に目を閉じ、刺激の停止後10分間もこの姿勢を維持し、外部の刺激に反応しなくなりました。
動物は眠っておらず、筋緊張は低下していませんでした。むしろ中程度の感覚刺激には反応しないようで、動きに関して不活発さが増加しているように見えました。
ネコをこの不活性な状態から目覚めさせるには、大きな音と、囲いを揺らすしかありませんでした。覚醒すると、ネコはミルクを飲むなどの刺激前の活動に戻りました。
コウベイシ博士はラットを使った実験で、前障の電気刺激によって意識障害が起きる事を裏付けました。(Bayat et al. 2018)
まず、ネズミは赤か緑の光に反応し、その色に応じた正しいポートに鼻を突っ込むことで餌のペレットをもらえることを学習しました。
ラットの前障に電気刺激を加えると、この課題の成功率が低下しました。これは刺激強度が強くなるほど顕著でした。
意識障害
刺激強度が高くなるほど、課題の成功率が減少した。
また約半数のラットで成功率が0%だったセッションが19回ありました。
これは前障刺激で誘発された不動状態、明らかな混乱、常同的な運動が原因でした。
一部のラットは最大5分間活動を停止し、セッションを完了できませんでした。これらのラットの行動は、前障刺激を受けた患者で観察された意識障害に似ていました。
誘発された常同的な運動は、首を反らし、肩を丸め、腹部を平らにし、体を後ろに引きずるという、一連の運動を特徴としました。
その後ラットは通常の姿勢に戻り、長時間の不活性な状態に陥りました。
運動停止
尾状核の電気刺激で、実行中の運動が停止することが示されています。人間を対象とした実験では、時に混乱を伴いました。
人間
ヴァン・ビューレン博士の研究
アメリカ国立衛生研究所のジョン・ヴァン ビューレン博士は、パーキンソン病の患者を対象に、尾状核頭部への電気刺激の実験を行いました。(Van Buren 1963)
患者の尾状核頭部に電気刺激を加えると、発話停止や運動停止が生じ、時に混乱を伴いました。
患者が数をかぞえたり、手を開閉している最中に刺激を受けると、それらの行動が止まりました。
その理由を尋ねると、たいていは刺激とは関係のない些細な言い訳をし、話したり動いたりする衝動が低下したという印象を残しました。
また、いくつかの症例では混乱した言動と、それに関する記憶の喪失がみられました。
58歳の男性患者E. B. 氏は尾状核の電気刺激を受けると、数かぞえを停止しました。
言い訳として「もう十分な所まで行ったと思ったから」と言いましたが、患者は止めと言われるまで数え続けるよう、事前に強調されていました。
その後も、刺激を受けるたびに突然の発話停止が観察されました。数え続けるように強く念を押されたにもにもかかわらず、彼は「何か障壁のようで、自然な終わり方だった」など、様々な言い訳をし続けました。
別のセッションでは、舌を突き出して保持し、右手をゆっくりと開閉し続けるように指示されました。電気刺激を受けると、彼は舌を引っ込め、右手の動作を止めました。
彼は右手の動作を止めたことは後で思い出しましたが、なぜ止めたのかは思い出せませんでした。
また別のセッションでは、彼は刺激をうけると最初の3秒間静かに横たわっていましたが、その後突然起き上がり、右側の検査官の方にわずかに向きを変えました。
最初、彼は反応がなく、シャツをまさぐり、話しかけられても面談者を見ませんでした。
その後、なぜ動いたのかと尋ねられると、「起き上がって話がしたかったんだ。何か伝えたかったんだ」と答えました。
あいまいに微笑んだ後、「大したことじゃない、ないよりはましな程度さ」と付け加え、ベッドに横になりました。
1分以内に彼は完全に意識を取り戻したようでしたが、発作中のどの部分も思い出せませんでした。
53歳の女性患者H. T. 氏も尾状核の電気刺激を受けると数かぞえを停止し、これは何度も再現されました。
数かぞえを止めた理由を尋ねられると、様々な答えが返ってきました。
例えば、「あなたは私に続けて欲しくないと思った」、「もう十分すぎるくらいやったと思った」 、「あなたが部屋を出ていくと思った」などです。
あるセッションでは、刺激を受けると彼女は頭を左に向け、喉でハミング音を発しました。また、目が素早く瞬きしていました。
刺激の後、彼女はどう感じたか尋ねられると、「看護師の一人が法廷でよく言う『Y、Z、オーイェー、オーイェー』って言っていました」と答えました。
看護師はその場におらず、隣接する場所からの声も聞こえませんでした。
58歳の男性患者L. S. 氏は尾状核の電気刺激を受けると混乱状態に陥りました。
静かに横たわっていましたが、刺激が始まると突然頭と目を右に向けました。
その後、彼は起き上がり、そうしないように言われたにもかかわらず、左手から検査用器具を取り外そうとしました。
数秒後、彼は言うことを聞くようになりましたが、言葉は発しませんでした。
その後すぐに彼は完全に意識を取り戻しましたが、発作中のどの部分も記憶に残っていませんでした。
東北大学の研究
日本の東北大学の研究者は、ヴァン ビューレン博士の研究でみられた人間の停止反応を再現しようと試みました。(Kwak et al. 1978)
被験者は疼痛患者とパーキンソン病の患者で、尾状核頭部に電気刺激を加えると、発話や自発的な運動を停止させることができました
また、まれに混乱が生じることがありました。
患者は数をかぞえるように、あるいは計算するように指示され、その作業中に電気刺激を加えられました。
すると患者は突然、数をかぞえるのを即座に、あるいは1~2秒以内に止めました。ほとんどの場合、刺激の解除直後、患者は妨害などなかったかのように、患者は計数を再開しました。
たとえば、ある患者が1から10まで数えるように言われました。
患者が数字の3に達したときに電気刺激が与えられると、刺激が解除されるまで計数が中断されました。
刺激が解除されると、患者は中断したところから数え直しました。
また、刺激によって計数が抑制されている間、患者はさまざまな質問を受け、ほとんど全ての事例で、刺激解除後に患者は刺激中に与えられた質問に正しく答えることができました。

さらに、ほとんど全ての事例で、患者は刺激の前中後に起こったすべての出来事を正確に思い出すことができました。
しかし、3つの事例では、刺激中に起こったすべての出来事を思い出すことができず、数かぞえを止めていることに気がつきませんでした。
このような場合、患者は刺激解除後に計数を再開しませんでした。このような混乱は刺激解除後すぐに、あるいは長くても1分以内に消失しました。
繰り返し刺激を与えると、患者は1から、または計数済みの数字から数え直したり、何かをつぶやいたり、時には停止反応が誘発されないこともありました。
刺激によって発話だけでなく、自発的な運動も停止しました。
例えば、ある患者に腕を持ち上げて、指を繰り返し曲げて伸ばすよう求めました。
尾状核に電気刺激を加えると、指の動作は停止しましたが、腕は持ち上げられたまま軽く揺れました。
刺激の解除後、患者は刺激前と同様に自発的な運動を行うことができました。
ネコ
ネコを使った動物実験でも、主に尾状核の電気刺激によって運動の停止反応が起きることが観察されています。
モントリオール神経科学研究所は、視床の電気刺激でネコの自発的な運動が停止することを発見しました。(Hunter and Jasper 1949)
歩いている時に刺激を受けたネコは、その場で凍り付いたように止まりました。

ネコは「呆然」というよりは「凝視」の表情で、じっと動かず、あるいはしゃがんだ姿勢のままでした。刺激がなくなるとすぐに、ネコは再び歩き始めました。
食事中であってもネコは突然動かなくなり、マウスを追いかけていた場合も、追跡中に停止しました。

光や音には反応しませんでした。目は開いたままで、目と口の周りがわずかにピクピクと動くことが多かったです。
脊髄反射と角膜反射は維持されていました。
刺激中に痛みに対して肢をひっこめましたが、激しい痛みに対してでさえ、ネコは防御行動をとりませんでした。
ブリティッシュコロンビア大学の研究者は、尾状核の電気刺激で、先行する実験に類似した運動停止反応をネコに誘発しました。(McLennan et al. 1964)
尾状核を10 Hz未満という低周波で電気刺激すると、ネコの自発的な運動が停止しました。
刺激が始まるとネコは即座に動きの途中で凍り付いてしまい、一歩踏み出す動作で片足を曲げたままにしてしまうほどでした。
しかし、尾がピクピクと動き続けることは頻繁に観察されました。
刺激期間の開始時にネコが食べ物に向かって前進していた場合、その注意はそらされず、目標をじっと見つめ続けました。
刺激が終了するとすぐに、ネコはためらうことなく前進しました。
刺激の周波数を高くすると、すべての動物で回転運動が生じました。
約10~15 Hzの周波数では、刺激の反対側を肩越しに見るかのように、ネコは頭を上げ、頭と首を回しました。
周波数をさらに上げると、動物は反対方向に回転し始め、回転速度は周波数に依存していました。
尾状核の電気刺激で学習した回避行動が抑制される、ということも示されています。(STEVENS 1961)
実験ではまず、ブザーが鳴ってから数秒後に与えられる電気ショックを避けるために、ネコは小さな開いた戸口を通過するように訓練されました。
続いて、ブザーが鳴る20秒前から電気刺激を尾状核に加えると、ネコはブザーが鳴っても、学習した回避行動をとらなくなりました。
ブザーが鳴ると、ネコは時々、戸口をくぐるかのように前足を上げましたが、その後立ち止って辺りを見回し、通過に失敗しました。
尾状核の電気刺激で食物を取得する運動が抑制される、ということも示されています。(Buchwald et al. 1961)
実験ではまず、ネコはレバーを押すと少量の液体クリームをもらえるということを学習しました。

尾状核に0.2 Hzの低周波で電気刺激を加えると、ネコのレバー押し反応はまず遅くなり、次いで停止しました。
刺激周波数を5 Hzまで上げると、効果が増大しました。刺激によってレバー押しが突然停止することもありました。
バーを押すのをやめた後、ネコはレバーを気にかけてないように見えましたが、フードカップに残っているクリームを舐め続けることはよくありました。
低周波刺激が終了すると、ネコは通常、束の間座った姿勢を保ち、続いて歩いて来てレバーを押し始めました。
低周波刺激によってレバーを押すのをやめたネコに300 Hzの高周波刺激を与えると、ネコはすぐにレバー押しに戻りました。
つまりネコのレバー押しは低周波刺激によってオフにでき、高周波刺激によってオンにできました。
その他の動物
サルなどの他の動物を使った実験でも、主に尾状核の電気刺激で運動の停止反応が起きる事が観察されています。
ロッチェスター大学の研究者は、前帯状皮質の電気刺激でサルの運動を停止させました。(Smith 1945)
アカゲザルの前帯状皮質に電気刺激を加えると、実行中の運動が直ちに停止し、サルの筋肉が弛緩しました。
薬物投与後に暴れているサルも、前帯状皮質に刺激を加えることによって完全に鎮静することができました。
刺激を受けた後、すべての動きが止まり、サルは目を閉じ、数分間、眠っているような静止状態のままでした。
パリ大学の研究者は、尾状核の電気刺激でサルの運動を停止させました。(Kitsikis 1968)
アカゲサルは拘束椅子に座った状態で、以下の図に示したさまざまな腕の運動を実行しました。
これらの運動はすべてブドウのかけらを得るために行われました。

尾状核の頭部に電気刺激を与えると、これらすべての運動を停止させることができました。
労力がかかる運動ほど、その停止は容易でした。
つまり、遠く高い所へ到達する動作や、またおもりを引っ張ったりする動作は、より弱い刺激で停止することができました。
また動作の開始を防ぐのは、動作自体を停止するよりも弱い刺激で済みました。
イヌを使った実験でも、尾状核の電気刺激によって運動の停止反応が起きることが示されています。(Goldzband et al. 1951)
尾状核への電気刺激は、イヌの自発的な歩行を停止させました
刺激が始まると、イヌは踏み出しを単純に完了させ、停止しました。
通常、尾を振るなどの他の定型的な動きも抑制されましたが、常にそうとは限りませんでした。
歩行停止に伴い、頭の振りむきや回転運動といった、いくつかの単純な動きが誘発されることがありました。
例えばある電極の刺激では、頭を左に向けることが歩行停止に常に伴いました。
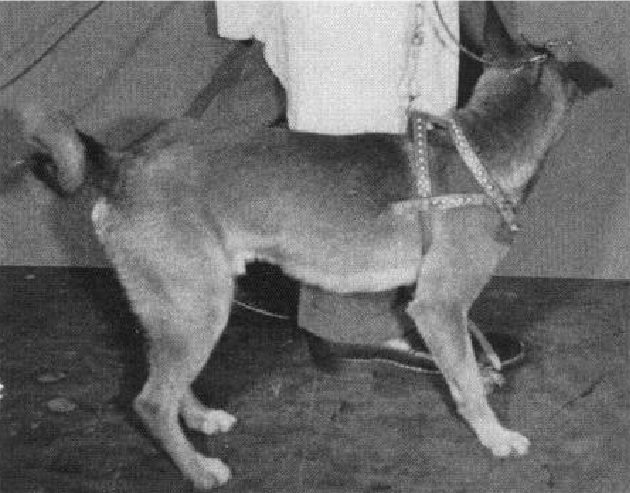
ウサギを使った実験では、尾状核の電気刺激によって全般的な行動の抑制が生じることが観察されています。(Klemm and Dreyfus 1975)
尾状核の電気刺激で、ウサギに以下のような行動への抑制効果が生じました。
- 箱に閉じ込められた時に、体をゆすることが少なくなった。
- 自由に行動させた時に移動量が減少した。
- ひっくり返された後に起き上がるまでの時間が長くなった。
- 嫌悪的な刺激である、風の吹きつけから逃げなくなった。
記憶喪失
海馬、扁桃体、そして尾状核の電気刺激により、ネズミに記憶喪失を誘発することが示されています。
海馬・扁桃体
頭皮から脳に電流を流して全身痙攣を引き起こす、電気痙攣ショックは記憶喪失を引き起こすことが知られています。(Kopp et al. 1966)
同様の効果を脳の特定部位への電気刺激によって誘発できることが、ネズミを使った実験で確認されています。
コロンビア大学の研究では、マウスの海馬への電気刺激が、電気痙攣ショックと同様の記憶喪失を誘発する事が発見されました。(Lidsky and Slotnick 1970)
まず、ケージの壁の一面に、触れると電気ショックを受ける金属棒を設置しました。
マウスにこのケージを探索させ、棒を触ると電気ショックを受けることをマウスに学習させました。
その後、マウスは海馬か新皮質領域への電気刺激、あるいは電気痙攣ショックを受けました。
翌日に再びケージを探索させると、海馬刺激を受けたマウスは、電気痙攣ショックを受けたマウスと同様の記憶障害の様相を呈しました。
つまり、マウスは金属棒に接触すると電気ショックを受けることを忘れてしまったようでした。
これは棒接触までの時間の減少、探索行動の増加 (下記の「垂直運動」と「活動」の増加)、凍り付き回数の減少で示されました。
記憶障害
扁桃体への電気刺激でもラットに記憶障害が起きる事が示されています。(Bresnahan and Routtenberg 1972)
まずケージの床を通電し、その上に台座を設置して、そこからラットが降りると電気ショックを受けるようにしました。
ラットをこの台座に乗せると、床に降りると電気ショックを受けることを即座に学習しました。
続いてラットが台座に戻った時に、扁桃体への電気刺激を与えました。
翌日、再度ラットを台座に乗せると、扁桃体への電気刺激を受けたラットは、そうでないラットに比べて床に降りるのが大幅に早くなりました。
ラットは床に降りると電気ショックを受けることを忘れてしまったようであり、記憶喪失が起きたと言えます。
また扁桃体のうち、内側核への刺激が最も有効でした。
記憶喪失
扁桃体刺激により、電気ショックを受けた後に床に降りるまでの時間が大幅に短くなった。
尾状核
ニューヨーク州立大学ストーニーブルック校の研究グループは、ラットの尾状核への電気刺激で、記憶障害が起きる事を発見しました。(Wyers et al. 1968)
ケージにレバーが設置され、ラットはそれを押すと甘い水を摂取することができました。
50回目のレバー押下時に、ラットは肢に電気ショックを受けました。
その後、0.1、1、5、30秒の遅延時間を空けて、尾状核への電気刺激を与えました。
翌日、ラットを再びケージに入れました。すると尾状核に刺激を受けたラットはほとんど躊躇することなく、レバーを押して甘い水を摂取しました。
飲水までに要する時間は、電気ショックを受ける前に測定したものとほとんど違いがありませんでした。
対照的に尾状核の刺激を受けてないラットは、レバーを非常に警戒しており、甘い水の摂取までの時間に大幅な遅れが生じました。
この尾状核による記憶障害は、電気ショックから脳刺激までの遅延時間によって大きな差はありませんでした。
記憶喪失
尾状核刺激により、飲水までに要する時間が大幅に短くなった。これは電気ショックから脳刺激の遅延時間によって大きな差はなかった。
同研究グループは続く実験で、尾状核刺激による記憶障害が、電気ショックから脳刺激までの遅延時間に依存することを示しました。(Wyers and Deadwyler 1971)
これは最大30秒から300秒と、前回の実験より遅延時間を大きくすることで達成されました。
実験ではラットにケージに設置した給水器から水を舐めさせ、50回目の舐め時にラットは肢に電気ショックを受けました。
その後、30、120、300秒という、前回より長い遅延時間を設けて、尾状核への電気刺激を与えました。
翌日、ラットを再びケージに入れると、尾状核刺激を受けたラットはほとんど躊躇することなく水を舐めるという、前回実験を裏付ける結果が得られました。
前回との違いは、電気ショックから脳刺激までの遅延が少ないほど、強力な記憶喪失がおきたことです。
記憶喪失
尾状核刺激により、飲水までに要する時間が大幅に短くなった。これは電気ショックから脳刺激までの遅延が少ないほど効果的であった。
尾状核への電気刺激でラットの学習を阻害するということも示されています。(Peeke and Herz 1971)
実験では、ラットはゴールに置いてある練り餌を目指してラシュリーIII 迷路を走りました。
1回の試行中にどの分岐点でも間違った方向に進まなければ成功となり、ラットは5回の試行のうち4回成功するまで迷路学習を行いました。
ラットはこの迷路学習中に、単発あるいは複数回の尾状核の電気刺激を受けました。刺激のタイミングはゴール到着後か、分岐点通過毎のいずれかでした。
するとラットの迷路学習が阻害され、基準達成までの試行回数が増加しました。特にゴール到着後にまとめて複数回の電気刺激を受けた場合が最も効果的でした。
記憶喪失
尾状核刺激により、ラットの迷路学習が阻害され、基準達成までの試行回数が増加した。
睡眠、無力症
尾状核、視床、前脳基底部の電気刺激で睡眠が誘発されることが示さています。
また前視床下部や尾状核の電気刺激で、無力症 (adynamia) と呼ばれる、筋緊張の喪失を伴う不活性な状態が生じることも示されています。
人間
尾状核
テュレーン大学の精神科医ロバート・ヒースは、アカゲザルと統合失調症の患者を対象にした実験で、尾状核頭部への電気刺激で睡眠を誘発できることを発見しました。(Heath and Hodes 2017)
アカゲザルでは、尾状核頭部に10 Hzで弱い電気刺激を与えると、サルは眠くなり、すぐに深い眠りに陥りました。
脳波パターンは通常の睡眠時に見られるものとそれほど変わらないものでした。
統合失調症患者の場合、尾状核頭部に100 Hzで2分間電気刺激を与えると眠気が起こり、急速に深い睡眠へと移行しました。
患者を覚醒させることは不可能でした。
患者の耳の近くで手を叩いても、最小限の反応しか誘発しませんでした。この反応は、刺激を止めてから数分間持続しました。
その後、徐々に覚醒が始まり、8~10分後には、刺激を受ける前の覚醒度に戻ったようでした。
視床下核
ソ連の科学者ナタリア・ベクテレワは、運動障害の患者を対象にした実験で、視床下核の電気刺激で眠気が誘発されることを示しました。(Bekhtereva 1969)
一人の患者には、15~30秒の間隔を挟んで視床下核への低周波 (4, 10 Hz) と高周波 (25, 50 Hz) の電気刺激が繰り返し与えられました。
すると覚醒と眠気が交互に現れました。
さっきまで活発だった患者が、眠くなり、あくびをし、眠気を訴え、うとうとし始めました。その後検査者が何もしなくても、患者はまた生き生きとして機敏になりました。
別の患者では、視床下核への高周波 (50 Hz) の電気刺激を15~30秒の間隔を挟んで4回繰り返すと、短時間の眠気が生じました。
ネコ
視床
スイスのチューリッヒ大学のヴァルター・ヘス博士は、ネコの視床の電気刺激で睡眠が誘発できることを発見しました。(Hess 1958)
4-6 Hzの低周波で視床間橋を電気刺激すると、ネコは体を落とし込んで、典型的な姿勢で眠りました。
叩かれたり揺すられたりすると、目が開くことが多かったです。
鼻の前に肉を置くと、ネコはすぐにそれを掴もうとしました。そして、食べ終わるとすぐにまた横になり、眠りに落ちました。
刺激を受けたネコは突然「気絶」するのではなく、快適な姿勢を探しました。何度か姿勢を調整しないとそれが見つからないこともありました。


前視床下部
前視床下部の電気刺激では、睡眠とは異なる不活発な状態が誘発され、ヘス博士はこれを無力症 (adynamia) と命名しました。(Hess 1958)
刺激を受けると血圧が低下し、ネコはその時点で維持していた姿勢から沈み込み、異常な姿勢をとりました。
骨格筋の緊張、自発的な活動、そして反応性は完全に失われ、実験者が触れてもネコは全く反応しませんでした。


尾状核
同じくチューリッヒ大学の別の研究者は、尾状核頭部の電気刺激でも無力症が生じる事を示しました。(AKERT and ANDERSSON 1951)
尾状核刺激後の数秒から2~3分で、それまで活発だったネコはおとなしくなり、座ったり横になったりし、絶えず目を細めました。自発的な活動は消失したように見えました。
外部刺激に対しては頭を回すだけで反応しました。それ以外は、疲れた不機嫌そうな表情をしており、活発な頭の姿勢とは非常に対照的でした。
全体的な様子からみると、ネコは完全に横たわり、いつでも丸くなって眠ってしまうのではないかと思われました。
しかし実際には、ネコは頭を上げたまま、座ったまたは横たわった姿勢で微動だにせず、あらゆる感覚刺激に対して反応しました。これは刺激が停止した後も数分間続きました。


前脳基底部の同期領域
UCLAの研究では、前脳基底部の同期領域に電気刺激を加えると、ネコに強力な睡眠作用が現れることが発見されました。 (Sterman and Clemente 1962, Sterman and Clemente 1962)
前脳基底部の同期領域とは、電気刺激によって大脳皮質に睡眠時に見られる脳波の同期活動を誘発できる領域です。
これは具体的には、ブローカ対角帯から外側視索前野に至る領域にあたりました。
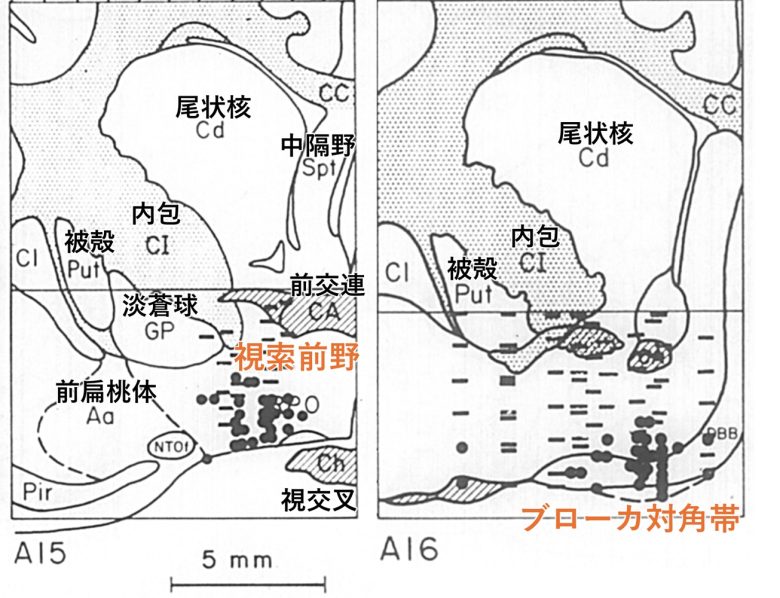
この領域は、ヘス博士が電気刺激によって無力症を誘発した前視床下部の領域の、やや前方に位置していました。
前脳基底部の同期領域に刺激を与えると、覚醒から眠気と睡眠への驚くほど急速な移行が起こりました。
刺激の開始によって始まる一連の現象は通常、進行中の行動の停止、続いてケージの隅への後退、そしてそこで自然なもたれかかる姿勢をとるというものでした。
これに続いて、目の前の環境への関心が明らかに失われました。
邪魔されなければ、もたれかかるネコは頭を床に垂らすか前肢の上に休ませ、目を閉じるか部分的に閉じ、見たところ眠りにつきました。
刺激開始から睡眠発生までの時間は、短くて5秒、長くて3分で、平均所要時間は約30秒でした。
邪魔せず放っておくと、ネコは眠り続けました。
また、前脳基底部の同期領域への電気刺激は、ラットへ攻撃中のネコを停止させ、そのまま眠らせるほど強力なものでした。
ダラウェア大学の研究により、前脳基底部の同期領域の電気刺激で睡眠が誘発されることが裏付けられました。(Lineberry and Siegel 1971)
実験ではまず、24時間ネコを絶食させ、その後にネコに食べ物を与えました。
ネコが餌を食べ始めて数秒後に前脳基底部の刺激を開始すると、ネコは食べるのを止め、餌皿から離れて横たわりました。
刺激を45~50秒間続けると、ネコは眠りに落ち、刺激を止めた後も眠り続けました。
サル
サルを使った実験でも、前視床下部の電気刺激で無力症 (筋緊張の喪失を伴う不活性な状態)が誘発されることが示されています。
前視床下部
コロラド大学の研究では前視床下部の電気刺激でアカゲザルに無力症が生じる事が示されました。(Sheer 1961)
前視床下部刺激により、サルははっきりとした静止状態になりました。
自由に動ける状況では、サルは不活性で、放っておくと概して静かに座り、時折目を閉じていました。
拘束ケージ内では、通常はサルは目を閉じ、居眠りしているように見えました。様々な刺激によって活性化しえましたが、反応は正常時よりも弱く、概して持続時間が短かったです。
刺激中のサルの行動の際立った特徴は、活動性の低下、刺激に対する反応性の低下、そして全般的な静止状態でした。
この行動は、瞳孔の収縮と全身の筋緊張の喪失を伴い、ヘス博士が無力症と表現した状態に似ていました。

イェール大学のホセ・デルガドも前視床下部の電気刺激でアカゲザルに無力症が生じる事を示しました。(Delgado 1964)
刺激を受けたサルは静止状態とうたたねを示しましたが、全身の筋緊張も失われており、この効果は自然な睡眠というよりはヘス博士が述べた無力症に似ていました。

拒食
尾状核や視床下部の電気刺激によって、拒食が生じることが確認されています。
デルガドの動物実験
アカゲザル
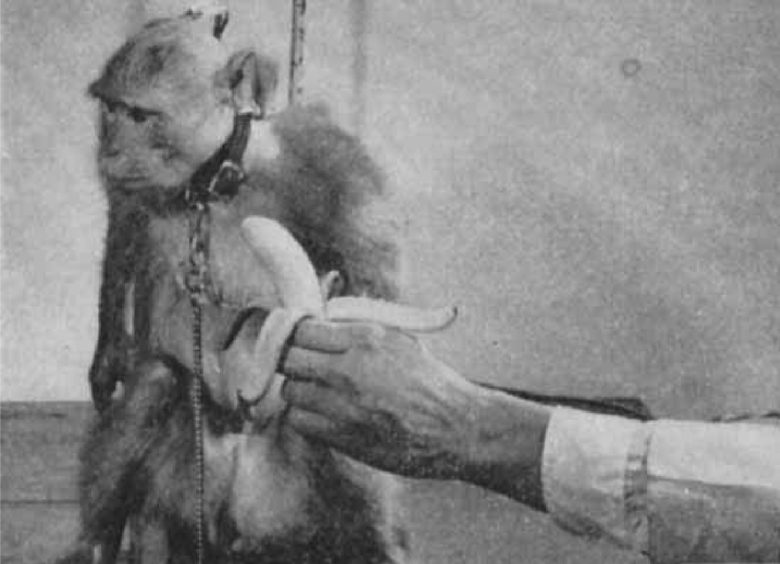
デルガドは尾状核頭部の電気刺激によってサルの食欲を抑制できることを示しました。(Rubinstein and Delgado 1963, Delgado 1964)
アカゲザルを椅子に拘束し、尾状核を電気刺激すると、食物に近づき、手に取り、食べるという行動が抑制されました。
果物が与えられると、サルは頭を向けて食べ物を見つめましたが、非常に近くにいてもそれを取ろうとはしませんでした。
この抑制効果は、ピーナッツ、ペレット、リンゴ、バナナ、オレンジなど、試したすべての種類の食べ物で観察されました。実験者がサルに食べさせようとしても、動物は食べ物から顔を背けました。
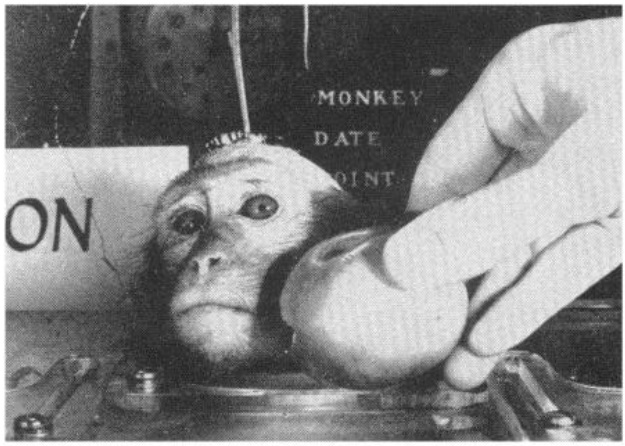
尾状核の刺激が始まると、咀嚼も停止しましたが、刺激が止まるとすぐに再開されました。
サルが餌に向かって歩いていた場合、刺激を受けると躊躇し立ち止まり、ゆっくりと離れていきました。
サルがすでにバナナを手か口に持っていた場合、刺激が始まるとすぐに果物はケージの床に落とされ、サルはそれ以上バナナに興味を示さず、他のサルがバナナを取ったときにも脅したり報復したりしませんでした。
サルは口の中に入っている食べ物を吐き出し、頬袋に食べ物がいっぱいになっている場合には、前肢を使って能動的に取り除こうとしました。
サルが水を飲んでいた場合、刺激が始まるとすぐに飲むのをやめ、ゆっくりと頭をそらし、飲用チューブから離れました。


テナガザル
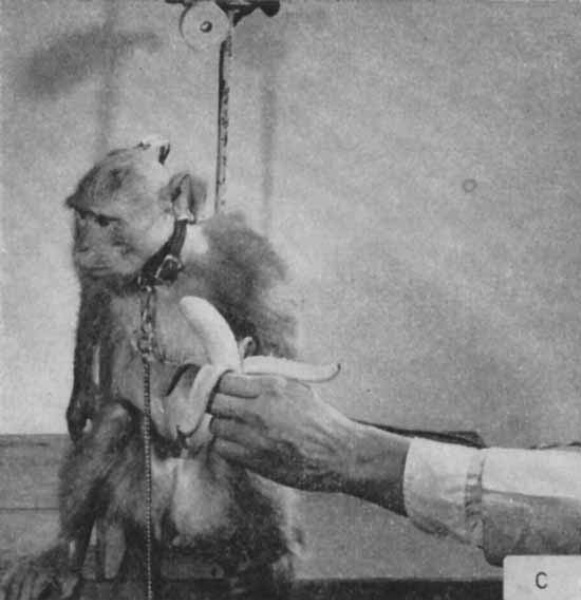
デルガドはテナガザルを使った実験でも、尾状核の電気刺激によって食欲を抑制できることを示しました。(Delgado 1975)
尾状核の頭部が刺激されると、リンゴを貪るように食べていた動作が即座に止まり、果実の欠片はゆっくりと口から引き抜かれました。
動物は刺激が続いている間は落ち着いており、刺激終了から数秒後に食事を再開しました。
また餌ペレットを得るためのレバー押し行動も尾状核刺激によって抑制されました。


その他の動物実験
尾状核
ダラウェア大学の研究では、尾状核への電気刺激でネコの食欲が抑制されることが示されました。(Lineberry and Siegel 1971)
実験ではまず、24時間ネコを絶食させ、その後にネコに食べ物を与えました。
尾状核を刺激すると、ネコは食べるのをやめ、餌皿の上で頭を静止したまま静かに立っていました。
ネコは凍り付いてはおらず、周囲を見回し、移動することができました。
内側視床下部
ペンシルベニア大学の研究者は、ラットを使った実験で、視床下部の内側部と外側部の電気刺激で摂食に関して相反する効果が生じることが示されました。(Smith 1961)
内側視床下部の電気刺激は摂食の抑制を引き起こし、これはラットが高度に欠乏した状態にあったときでさえ生じました。
特に、絶食後に食物が与えられた最初の30分間で、この抑制は非常に顕著でした。
対照的に、外側視床下部の電気刺激は摂食の促進を誘発し、識別能力を欠いた摂食または咀嚼が生じました。
動物は、食べ物、木片、排泄物、ペーパークリップなど、偶然出会ったものは何でもつかみ、口に入れ、噛み続けました。
その後、ラットはそれらの物体を落として、別のものを掴み、刺激が終わるまでその行動を続けました。
テネシー大学の研究者は、ラットを使った実験で同様の観察結果を得ました。(Morgane 1961)
内側視床下部の電気刺激は摂食を抑制し、空腹のラットでさえ刺激を受けると摂食を停止しました。
対照的に、外側視床下部の電気刺激では摂食を促進し、満腹のラットでさえ刺激を受けると摂食を開始しました。
食欲を促す外側視床下部と、食欲を抑える内側視床下部の同時刺激を行うと、双方の効果が相殺されました。
