性行動

人間の脳の特定部位の電気刺激により、性的な快楽と欲求、勃起、オーガズム、恋愛感情などを引き起こすことができます。
このような部位には中隔野、扁桃体、視床下核、視床などがあります。
動物の脳の特定部位の電気刺激により、交尾、勃起、自慰行為、射精、排卵、子宮収縮 (加えて出産・流産) などを引き起こすことができます。
このような部位には中隔野、扁桃体、視索前野、内側前脳束、腹側被蓋野、被殻、水道周囲灰白質、視床下部、下垂体などがあります。
これを悪用すると、そのような傾向のない個人に性犯罪を犯させることも可能になると思われます。
目次All_Pages
回路
オスの性行動
オスの性行動に関与する領域には、内側視索前野、外側中隔、内側扁桃体、内側前脳束、腹側被蓋野などがあります。
内側視索前野はオスの性行動の重要な統合部位で、性的な動機付けと交尾の双方を促進します。(Hull and Rodríguez-Manzo 2017)
外側中隔もオスの性行動に重要な部位で、性的な満足感と動機付けの両方に関与しています。 (He et al. 2013)
また勃起時や射精時に活性化し、またテストステロンの受容体が豊富に存在する部位でもあります。 (Gulia et al. 2018, Mallick et al. 2007, Menon et al. 2022)
外側中隔から内側視索前野へは、内側前脳束を経由したつながりがあります。(Swanson and Cowan 1979)
内側扁桃体は内側視索前野と外側中隔の上位にある部位で、オスの性行動の制御に関与しています。(Jennings and de Lecea 2020, Hull et al. 2002, Caffé et al. 1987)
腹側被蓋野は前ページで説明の通り、欲求に関して重要なドーパミンニューロンが存在する部位です。
内側視索前野、外側中隔ともに、外側視床下部の内側前脳束を経由して腹側被蓋野を活性化し、オスの性行動を促進します。(Bayless et al. 2023, Sheehan et al. 2004, Conrad and Pfaff 1976)
以上で説明した領域の電気刺激は、オスの性行動を促進することが示されています。
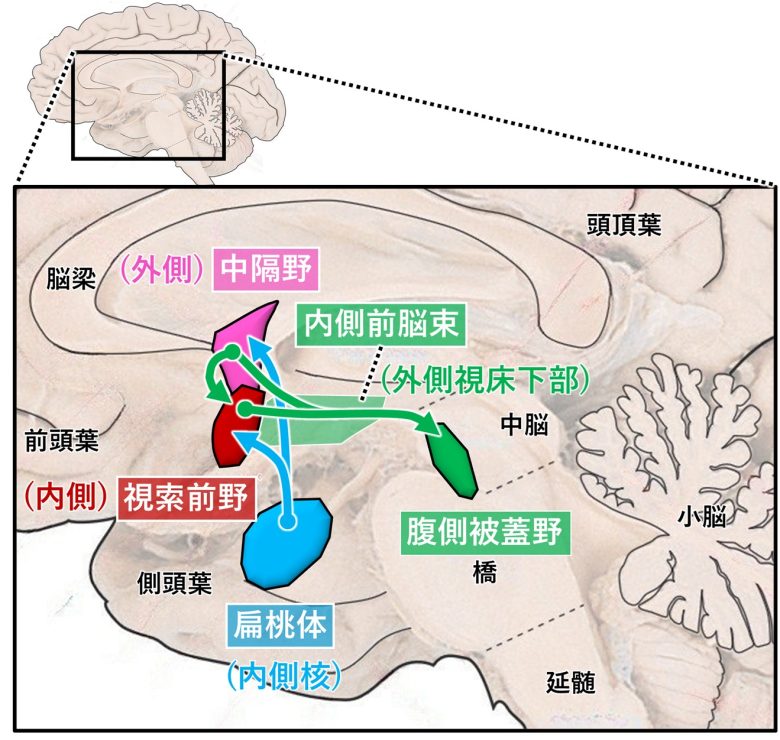
オスの性行動に関与する領域、内側扁桃体、内側視索前野、外側中隔、内側前脳束、腹側被蓋野
(脳画像はVanderah 2018から)
メスの性行動
メスの性行動では、視床下部の腹内側核がメスの性行動を制御するための最も重要な脳領域です。(Yin and Lin 2023)
視床下部の腹内側核を活性化するとメスの性受容性を迅速に高めることができ、逆にこの領域の損傷はそれを抑制します。
視床下部の腹内側核は攻撃行動が生じる領域でもありますが、性行動と攻撃行動は地理的に異なる小領域に細分化されています。
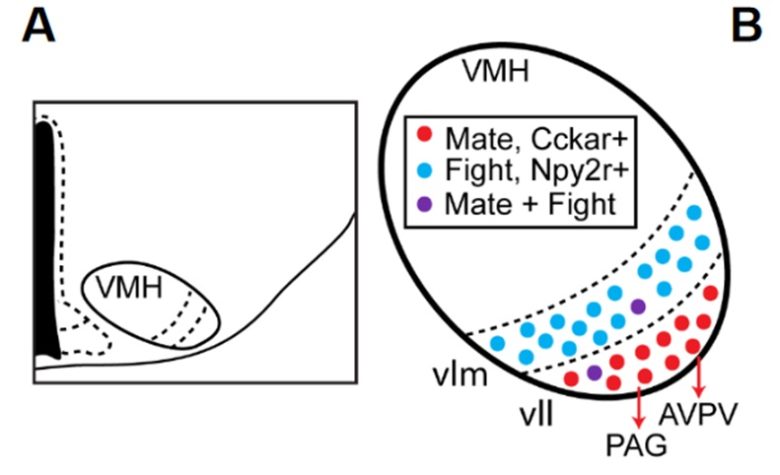
そして視床下部の腹内側核の電気刺激で、メスの性行動が促進されることが示されています。
人間
中隔野、扁桃体、視床下核、視床などの電気刺激で性的な快楽・欲求が生じることが示されており、時にオーガズムに至ることすらあります。
誘発される反応は制御できないほど強くなることもあり、これは例えば、このような脳部位を強迫的に自己刺激する患者に表れます。
また側頭葉、扁桃体への電気刺激で恋愛感情のようなものが生じる事が観察されています。
ヒースの人体実験
性的な快楽・欲求
テュレーン大学の精神科医ロバート・ヒースは、主に統合失調症の患者を対象に、脳電気刺激の人体実験を繰り返していました。(Heath and Hodes 1954, Heath 1964)
患者は主に中隔野に電気刺激をうけましたが、結局のところ統合失調症への治療効果はなかったと結論づけられました。
注目すべき点は、中隔野への電気刺激で快楽が生じ、程度の差はあれ、患者は一貫して性的に動機付けられたことです。
例えば、ある患者は父親の命に関わる病気について語って泣きそうになり、自分にも何らかの責任があると非難していました。
しかし中隔野が刺激されると、すぐにこの会話を打ち切り、15秒以内に、女友達とデートして誘惑する計画について語り、ニヤニヤした満面の笑みを浮かべました。
なぜこんなに急に会話を変えたのかと尋ねると、彼はその女の子に関する計画が突然思い浮かんだと答えました。この現象は刺激を繰り返すたびに再現しました。
別の患者は、2年以上にわたってひどく焦燥し、落ち込み、自己非難と絶望感を表明していました。
しかし、中隔野が刺激されると、彼は大きな笑顔をみせ、1分もしないうちに若い頃の性体験を語りました。
また3人の男性患者は、中隔野の刺激中に陰茎勃起を経験しました。
一方、外側視床下部や中脳被蓋などに刺激を加えると、「これは良い気分だ」といった快楽の申告がありましたが、患者は性的には動機付けられませんでした。
自己刺激
28歳のナルコレプシーの男性患者「B-7」には、中隔野、海馬、中脳に電極が取り付けられ、携帯型の刺激装置でこれらの部位を自己刺激できるようにしました。(HEATH 1963)
患者はこの刺激装置を17週間装着しましたが、この期間中でほとんどもっぱら、中隔ボタンを押していました。
なぜ中隔野のボタンを高い頻度で押すのか聞くと、患者は「気持ちがいい」と述べ、まるで性的なオーガズムに向かっているかのようだったと言いました。
しかし、オーガズムの絶頂に達することはできなかったと申告し、ボタンを頻繁に、時には狂乱的に押すのは絶頂に達するための試みであったと説明しました。
さらに中隔刺激には覚醒効果があり、患者はこれでナルコレプシーと闘うことができました。
25歳のてんかんの男性患者「B-10」は、合計17の異なる脳領域を自己刺激しました。(HEATH 1963)
これらの領域は無作為に選択され、装置の設計上、一度に3つの領域を接続することができました。
最も頻繁に押されたのは、視床中心正中核を刺激するボタンでした。しかし、この刺激は快い反応を誘発するものではなく、むしろイライラを誘発しました。
患者の申告によれば、この刺激中に記憶を思い出しそうになったが、よくつかめなかったとのことでした。頻繁な自己刺激は、このとらえどころのない記憶をはっきりさせるための努力でした。
患者は、中隔野の2つの電極と中脳被蓋の1つの電極への刺激で最も一貫して快感を申告しました。
快感反応とともに、中隔刺激によって患者に性的な思考が頻繁が生じました。実際の内容は様々でしたが、元の感情状態や部屋で交わされていた話題には関係がありませんでした。
患者はニヤニヤした満面の笑みを浮かべ、思い浮かんだ性的な思考を会話に持ち出しました。
このことについて質問されると、彼は「なぜそんなことが頭に浮かんだのかはわからない。たまたま思いついただけだ。」と答えました。
一方、中脳被蓋の刺激では「幸福感」が生じましたが、性的な思考は伴いませんでした。
彼はまた、扁桃体と尾状核の2つの部位への刺激を「良い」としましたが、快感の質はやや劣りました。
患者は、視床正中核への刺激によって生じた欲求不満や怒りが、中隔野と中脳被蓋への刺激によって緩和させることができたことに気づきました。
本人に告知せずに中隔野を刺激したときは、ほぼ瞬時に、彼の行動状態は普段の無秩序、激怒、迫害的なものから、幸福で穏やかな多幸感を伴うものへと変化しました。
オーガズム
34歳の女性のてんかん患者「B-5」には電気刺激ではなく、アセチルコリンとノルアドレナリンによる中隔野への化学刺激が行われました。 (HEATH 1972)
注入終了後から1~2分で、患者の気分は徐々に変化し始めました。徐々に気分が高揚し、10分から15分以内に、意識の高まりを伴う穏やかな多幸感に至りました。
変化の程度は、刺激を与えたときの患者の状態に左右されました。
気分が沈んでいた場合は劇的な変化を示し、処置の開始時にすでに気分が良かった場合は、変化はそこまで著明ではありませんでした。
気分の高揚と意識の高まりは、性欲の増加を伴い、ほとんどの場合、5分から10分以内に反復的なオーガズムで最高潮に達しました。
患者は質問されたときにその反応を説明しただけでなく、彼女の官能的な外見と動きからも確認することができました。
同性愛者の転向実験
1972年、ヒースは脳電気刺激によって同性愛者の男性を異性愛者に転向させるという、悪名高い実験を行いました。(HEATH 1972)
ヒースはこの「治療」に成功したと主張していました。
被験者はてんかんを患う24歳の男性「B-19」で、5年間の同性愛歴と3年間の薬物乱用歴がありました。
「毎日自殺を考えて生きている」と繰り返し発言し、何度かの自殺未遂がありました。
この患者は異性間性交をしたことがなく、異性間性交のビデオを実験室で患者に見せると、彼はビデオに嫌悪感を示し、怒って抗議しました。
そして翌日から中隔野への、実験者からの受動刺激および自己刺激の両方を含む「治療」プログラムが開始されました。(Moan and Heath 1972)
中隔野の受動刺激の3回の期間中と終了後、被験者は気分が改善し、頻繁に笑い、物事をはっきりと考えられるようになったと述べ、全身の筋肉が緩んだ感覚を申告しました。
彼はこれらの反応を、アンフェタミンの使用を通じて求め、経験した快楽状態に例えました。
さらに、彼は性欲が増加していることを申告しました。
次の2回は患者に中隔野を自己刺激させました。
この自己刺激のセッション中、被験者は圧倒的な多幸感と高揚感を経験しました。彼の激しい抗議にもかかわらず、接続を切断せざるをえませんでした。
次の4日間、被験者の気質と行動に顕著な改善が見られました。
研究室でも病棟でも反抗的でなくなり、より協力的になり、女性職員への関心が高まりました。また彼は自慰行為への衝動を伴う、性的な興奮を経験しました。
翌日の午後、彼はためらうことなく異性間性交のビデオを再度見ることに同意しました。上映中に彼は性的に興奮し、勃起し、自慰行為でオーガズムに達しました。
このセッションの終わりに被験者は「気分がいい」と述べ、非常に満足していました。
その後の4日間にわたる被験者の行動は、自己満足度の増大、性行為への執着、そして女性に対する関心の継続的な成長を示しました。
中隔野への受動的な刺激が再開されると、被験者は覚醒感、高揚感、そして非常に「ハイ」な状態にあると報告しました。その後の自己刺激でも、温かさ、紅潮、性的興奮を経験しました。
この時点で、患者は女性に対して積極的な関心を持ち続けており、近い将来に異性愛行為を試みたいという願望を表明していました。
そして、彼は売春婦を紹介され、完全なプライバシーを確保するために特別に準備された研究室で2時間の性行為に及びました。
不自然な状況と、深部脳波を測定するためのリード線につながれていたも関わらず、被験者は積極的に性交に参加して挿入に成功し、非常に満足のいくオーガズムが得らました。
患者はプログラムの終了後、彼は既婚女性と出会い、ほぼ10か月間親密な性的関係を築きました。
同性愛的な行動は、お金が必要で、仕事がないときに、2回だけあったと報告されました。
デルガドの人体実験
恋愛感情
イェール大学の神経生理学者ホセ・デルガドも、統合失調症の患者やてんかん患者を対象に脳電気刺激の人体実験を行っていました。(Delgado and Hamlin 1960)
彼は複数のてんかん患者の側頭葉への電気刺激で快楽が生じたことを示しました。
3人の患者では、快感に加えて恋愛感情のようなもの、具体的には、初対面の男性面談者への好意と結婚願望が誘発されました。
1人目は36歳の女性患者で、右側の側頭葉の表面から3 cm下の所の刺激で、顔から足の先までにおよぶ左半身に、快いうずく感覚が誘発されました。(Delgado 1970)
彼女はクスクス笑い、おかしな発言をし始めました。彼女はこの感覚を大変気に入りました。
刺激を繰り返すと彼女はより話し好きに、誘いかけるような態度になり、初対面の男性面談者と結婚したいという彼女の欲求を明け透けに表明しました。
刺激の前後の面談では、彼女の態度はかなりしっかりしており、親密さや過度の人懐っこさはみられませんでした。
2人目は30歳の女性患者で、扁桃体への電気刺激はくつろいだ、快い感覚を誘発しました。
彼女の発言量は顕著に増加し、それはより親密な性質を帯びるようになりました。
そして初対面の面談者への好意を率直に表明し、彼の手にキスをして、今彼女にしてくれていることへの計り知れない感謝の念について語りました。
別の日に扁桃体の同じ地点を刺激すると、似たような発言量の増加と感情の状態が再現されました。
刺激のない面談では、彼女はかなり控えめで落ち着いた様子でした。
3人目は女性への帰属傾向がやや強い、11歳の少年の患者でした。(HIGGINS 1956, Delgado 1970)
例えば、彼は刺激前の面談では、男性面談者に対して「香水の匂いがするね、可愛い」などと言ったり、面接官の匂いを嗅ぎ、舐め、キスし、噛んだりしました。
面談中に左側頭葉の表面の地点を刺激すると、唐突で率直な快楽の申告がありました。
患者は刺激前は黙り込んでいましたが、この刺激後、彼は「ねえ、これをくれるなら僕をもっと長くここにおいておいてもいいよ!これ好きだよ」と叫びました。
彼は続けて、この脳のテストはいい気分にさせてくれると言いました。
合計16回の同地点の刺激のうち、8回の刺激は「気持ちいい 」という力強い表現とともに、どれも似かよった発言を引き出しました。
これらの発言のうちいくつかは、男性面談者への好意を含んでおり、最後のものは少年の官能的な伸びを伴っていました。
これらの反応は刺激前の面談では見られませんでした。
別の面談では、同地点の刺激後、彼は自身の性同一性について戸惑いを見せました。
そして彼は突然、結婚したいという彼の欲求について語り始めました。
「誰と?」と聞くと、彼は即座には答えませんでしたが、1分20秒の沈黙の後、患者は「考えたけど——あれは——あなた (男性面談者) に言っていた思う」と言いました。
そしてこの話題は完全に打ち切られました。
続く40分間では他の地点がテストされましたが、患者はまったく異なる無関係な内容の、いくつかの話題について語りました。
再び件の地点が刺激されると、彼は自身の性同一性について再度疑問を呈し、「僕は少年なのか少女なのか、どっちになりたいんだろう、と考えてたんだ」と言いました。
さらなる刺激の後、彼は明確な喜びとともに「今やってるね」と発言し、さらに「僕は少女になりたい」と言いました。
セム=ヤコブセンの人体実験
性的快楽
ノルウェーの神経生理学者セム=ヤコブセンも統合失調症やパーキンソン病の患者を対象に脳電気刺激の人体実験を繰り返していました。(Sem-Jacobsen 1968)
前脳の腹内側部や大脳辺縁系の電気刺激により、幸福感や多幸感といった、肯定的な気分の変化が生じました。
誘発された肯定的な反応は以下のように3種類に分類されました。
- 患者がくつろぎ、安心し、幸福感を感じ、少し眠くなることがある。
- 患者は確実に変化し、機嫌がよく、気分がよい。患者はくつろぎ、安心し、楽しんでいる。笑顔が多い。わずかな多幸感があるが、行動は正常範囲内。彼はもっと刺激を欲しがるかもしれない。
- 多幸感は明らかに正常範囲を超えている。患者は大声で笑い、楽しみ、肯定的に刺激を好み、もっと刺激を欲しがる。
明確にそれとわかる性的快楽は、2人の男性に生じました。
1人の患者は強い性的快楽を得ました。
患者は電気刺激を好み、再度刺激を受けたいと言いましたが、彼が突然満足してしまうと、それ以上の電気刺激を望まなくなりました。
ある日、彼は自発的に以下のような情報を提供しました。
- くつろいでいる。
- 気持ちいい。
- 性的快楽のようだ。
- 臭いはない。
- 味もない。
- 全身で感じます。
これが間違いなく性的な反応であることがわかると、それ以上の刺激は行われませんでした。
もう1人の患者は、かなり静かで無口な男性でした。彼は刺激が気に入ったようで、もっと刺激を求めてきました。彼の反応は以下の通りでした。
- 頭を動かす。
- 深呼吸。
- ベッドの上で動き回る。
- 脚を動かすー紅潮ー絶え間ない震えー深呼吸。
- 突然の弛緩ー笑顔。
- 動きが止まるー紅潮ーベッドを濡らす。
- 射精。
- 後に目を潤ませ、周りを見渡す。
他にも何人かの患者にオーガズムのような反応がみられました。
自己刺激
セム=ヤコブセンもヒースと同様、自分の患者に脳を自己刺激をさせました。(Sem-Jacobsen and Torkildsen 1960)
患者が自分自身を刺激する方法は、部位によって異なりました。
ある部位では、患者は刺激を長時間続け、短い休憩のみで中断するのを好みました。他の部位では、頻繁に刺激の開始と停止を繰り返すことで最大の快感を得ているようでした。
強い快感部位では、患者は自分自身を刺激して痙攣の発作を起こすことがわかりました。
発作後には、これらの患者はくつろいで横たわり、幸せそうに笑っていました。
ベクテレワの人体実験
性的な快楽・欲求
ソ連の神経科学者で心理学者のナタリア・ベクテレワは、パーキンソン病などの運動過多症の患者を対象に脳の電気凝固術を施していました。(Bekhtereva 1969)
その術前には脳の電気刺激の実験が行われ、視床、視床下核、中脳被蓋の電気刺激で性的な快楽・欲求が生じることを発見しました。
37歳の女性患者はパーキンソン病を患っていました。
視床の外側腹側核とその隣接部位を電気刺激すると、患者は極度の快感を伴う性的興奮を経験し、それはオーガズムに変わりました。
これは刺激を停止すると消失しました。
官能的な体験には特徴的な表情や自律神経の変化が伴いました。
患者はより頻繁に研究室を訪れるようになり、そこでスタッフと会話を始めました。また、病棟の入り口、廊下、または病院の庭で彼らを待ち、次の電気刺激がいつ行われるのかを聞き出そうとしました。
彼女は処置の回数を増やすように要求し、延期されると不満を述べ、研究室に行くように言われるまでイライラした様子を見せました。
患者は刺激を繰り返し受ける方法を積極的に探していました。
彼女は電気刺激を施す人物に特に注意を向けました。この人物に対して、患者はある種の性的な色合いをもった奇妙な熱狂を抱くようになりました。
性的な衝動を彼女は恥ずかしがり、それを他人に知られたくなく、自分の行動に細心の注意を払い、衝動の発現を隠し、気を紛らわせ、何らかの活動に集中しようとしました。(他の患者の世話を積極的に手伝い、読書や音楽を聴くなど)
49歳の女性患者はパーキンソン病を患っていました。
左の中脳被蓋、視床下核、視床の外側腹側核を電気刺激すると、患者に快い感覚が生じました。
これは右半身、特に腕の異常な温感を伴う、並外れた快感でした。これらの感覚の強さは、刺激の強さが高いほど強くなりました。
視床の外側腹側核の刺激では振戦も止まり、硬直も軽減しました。
刺激終了と同時にこの現象は消失しました。
患者の衝動は電気刺激を受けた後の最初の数時間が最も顕著で、患者の気分は楽しく、高揚していました。
彼女は男性と冗談を言ったり、いちゃついたりするようになり、時にはあきらかに性的な性質を帯びた、あいまいな言葉で話しかけました。
彼女は刺激を繰り返すように執拗に要求し、また快感を味わいたいと説明しました。
患者は自分に注意を向けさせようと努力し、自分をきれいに見せ、唇をすぼめたりしました。
次第にこれらの発現は軽減されていきましたが、数日後、患者は医師たちに迫り、 「快楽電極」による刺激を繰り返すよう求めました。
そして次の電気刺激で衝動が強まり、同じ特徴的な行動パターンがよみがえりました。
刺激中の快楽が強ければ強いほど、衝動の発現はより顕著になり、より長く続きました。
その他の研究
性的な体験的現象
モントリオール神経科学研究所のピエール・グローア博士は、扁桃体の電気刺激で性的な体験的現象が生じた症例を報告しました。(Gloor 1986)
対象は40歳のてんかんの女性患者で、反応は右扁桃体刺激によって生じました。患者は困惑した様子で、自分が感じたことについて話したがりませんでした。
彼女は吐き気のような腹部の感覚を覚えましたが、それに加えて、外陰部と太ももの内側に、まるで性交をしているかのように快感を覚えました。
彼女はパートナーの姿は見ませんでしたが、それが「X」であることは分かっていました。Xとは、彼女が初めて性交をし、その後も何度も性交を重ねたボーイフレンドでした。
彼女は、その体験は古い記憶の想起であると断言し、その後も他の男性と性交したにもかかわらず、扁桃体刺激によって想起された感覚は、昔の恋人「X」との性交時の感覚であると確信していました。
自己刺激
アルベルト・アインシュタイン医学校では、48歳のヘルニア疼痛の女性患者に、治療として視床の後外側腹側核への電気刺激が施されました。(Portenoy et al. 1986)
患者には自己刺激装置が手渡され、刺激はある程度の鎮痛をもたらしました。
しかし患者は、刺激によって性的な快楽も同時に得られることに気づきました。この快楽反応は、連続的な刺激によって高まりました。
患者は性的に著しく興奮しましたが、オーガズムは生じませんでした。
不整脈や神経症状が生じましたが、患者は刺激装置を強迫的に使用するようになりました。
最も頻度が高いときには、患者は一日中自己刺激を行い、自身の衛生や家族の用事を無視しました。
振幅ダイヤルを調節する指の先端に慢性的な潰瘍ができ、刺激の振幅を大きくしようと頻繁に装置をいじりました。
時には、刺激装置へのアクセスを制限するよう家族に懇願し、その都度、短時間の休止の後に刺激装置を戻すよう要求しました。
性的な快楽・欲求
インドの研究者は、パーキンソン病の治療として、視床下核への電気刺激で2人の患者で性欲が過剰になった症例を報告しました。 (Bhargava and Doshi 2008)
1人目は70歳の男性患者で、電気刺激を開始してから1か月後、彼の妻は彼が性欲過剰になったと述べました。
彼は毎晩性的な満足を求め、拒否されると非常に攻撃的になりました。満足すると、彼は通常の自分に戻りました。
これは患者にとって疲れることでしたが、彼は衝動を制御できないことに気づきました。
彼の性欲過剰行動は手術後ほぼ4年間続き、その後、介入なしに突然停止しました。彼の妻は、この変化は彼が最近宗教指導者と交流したことによるものとしました。
二人目は58歳の女性患者で、術後、患者は夫の部屋に無理やり入り込んで性行為を要求するようになりました。
さらに、患者は自制心を失い、家族の他の男性にも自分をさらして性行為を要求しました。
術前、時折の例外を除き、数年間夫と性的関係を持っていませんでした。
術後、彼女は世話をする看護師に性欲や欲求不満を声に出して表現するようになりました。
また患者は甘いものが好きでしたが糖尿病で、この渇望が術後に増加しました。
運動障害の治療効果は素晴らしいものでしたが、異常な行動は5年間続き、抗精神病薬による介入が必要でした。
中国の研究者は、強迫性障害の28歳の男性患者に、治療として内包前脚への電気刺激を施しました。 (Chang et al. 2010)
テスト刺激では患者は軽度の多幸感と強迫観念の減少を申告しました。これらの感情は、刺激をオフにすると急速に消えました。
継続的な電気刺激が3 Vで開始され、それから約3週間後、彼は活力がみなぎり、睡眠欲求が減少し、性欲が高まり、性行為を頻繁に考えるようなったとを申告しました。しかし強迫症状は持続しました。
患者は保守的なキリスト教徒で、性欲の増加に不快感を覚えていると述べました。
強迫症状の緩和を試み、刺激強度が4 Vに増加されました。
2週間後の診察で、彼は気分が急激に変わり、抑えきれない性欲が彼をパブの女性との一夜限りの関係に駆り立てたと申告しました。 彼の母親は、彼が以前よりも怒りやすくなったと申告しました。
彼の強迫性障害の症状が持続したため、さらに刺激強度が4.5 Vに増加されました。
しかし数日後、彼は持続する症状と性欲過剰により、重度の自殺念慮に苦しみました。
彼は入院し、刺激強度が1 Vに減少すると彼の性欲過剰は治まりました。残念ながら状態は手術前と同じだったため、患者は落ち込んでいました。
勃起の感覚
スタンフォード大学の研究者は、前帯状皮質の電気刺激で性的興奮の他、勃起感覚が誘発された症例を報告しました。(Yih et al. 2019)
実験では18人のてんかん患者を対象に、前帯状皮質、島皮質、眼窩前頭皮質への電気刺激で誘発される主観的体験や感情状態が調査されました。
すると男性被験者2名に、前帯状皮質への電気刺激で性的興奮が生じました。
一人目の被験者は、4 mAの刺激で「性的に興奮した」感覚を申告し、6 mAの刺激では、「エロチックな」感覚はより強くなり「おお!」と驚きの声を上げました。
別の地点の刺激では、実際には勃起は伴わない、勃起の感覚が生じました。
二人目の被験者は、5 mAの電気刺激で「非常に性的に興奮した」感覚を申告しました。この部位への刺激は、内臓感覚や皮膚感覚も誘発しました。
オス
動物のオスでは、中隔野、内側視索前野、内側前脳束、腹側被蓋野、内側扁桃体の電気刺激で、勃起、射精、自慰行為、交尾など、様々な性行動が誘発・促進されることが示されています。
勃起・射精
中隔野・視索前野
アメリカ国立衛生研究所は、リスザルの脳内で勃起を誘発する領域を広範囲に探索しました。(MacLean and Ploog 1962)
すると中隔野と内側視索前野の電気刺激で勃起が生じることがわかりました。
20 -30 Hzの電気刺激が最も効果的で、刺激から3 -6秒以内に勃起が即時に生じました。
内側前脳束と視床下核と腹側被蓋野からも勃起が得られ、こちらは60 -100 Hzの刺激が効果的でした。
さらに、勃起には排尿、発声、不整脈が伴うこともありました。
ヤーキーズ霊長類研究所は、視索前野の勃起誘発部位を特定のパターンで繰り返し電気刺激することで、アカゲザルの射精を誘発することに成功しました。(Robinson and Mishkin 1966)
精液には運動性精子が高濃度で含まれていました。
誘発された射精は、いくつかの重要な点で正常な反応に似ていました。つまり、射精には完全な陰茎勃起が先行かつ伴い、また腰を突き出す動作を伴い、精液の放出は射出性でした。
射精後、動物は通常は静かに座り、しばしば居眠りをしていました。
最も効果的だったのは、それぞれが10~20秒間続き、20~60秒の間隔を空けた一連の刺激列でした。
この方法で刺激列を適用すると、勃起は徐々に大きくなり、腰の突き出しを伴う射精が5~10回目の刺激で起きました。
20 Hz未満または100 Hzを超える周波数では勃起は誘発されず、射精につながる大きな勃起は40~50 Hzでのみ発生しました。
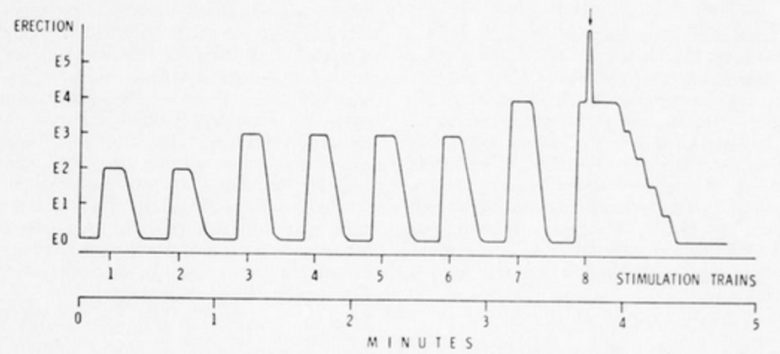
1匹のサルに、レバーを用いて同じ脳部位を自己刺激させました。
2~5分間の休止を挟んで8~10連の連打を繰り返すことで、大きな勃起が誘発され、8回目の連打でサルは腰を突き出して射精に至りました。
その後、レバー押しは完全に停止しました。
全体の動作は25分間続き、その間に記録されたレバー押しはわずか76回でした。しかし、この現象が他の2回の機会でも繰り返されたことから、非常に低い速度ではあったものの、サルは特定の時間パターンで確かに自己刺激を行っていたことが示唆されました。
インドと日本の共同研究では、勃起における中隔野の役割が調査されました。(Gulia et al. 2008)
中隔野への電気刺激がラットの覚醒時とレム睡眠時に行われ、いずれの場合でも勃起が生じました。
ほとんどの場合で自発的なものと同じである完全な勃起が生じましたが、一部で排尿を伴う異常な勃起も生じました。
レム睡眠中の刺激では、1回の3秒間の刺激で最大16分続く、持続型の勃起が得られることもありました。
勃起は外側中隔からは効果的に誘発されましたが、内側中隔からは誘発されませんでした。
被殻
ヤーキーズ霊長類研究所はアカゲザルを使って脳内で勃起を誘発する部位を探索しました。(Robinson and Mishkin 1968)
勃誘発部位は多くが視索前野にありましたが、意外なことに、被殻と前帯状皮質においても勃起誘発部位が発見されました。
また、視床下部や扁桃体にもいくつか勃起を誘発する部位が見つかりました。
内側前脳束
オックスフォード大学の研究者は、脳の自己刺激の実験の最中に、2匹のラットが精液を放出していることに気づきました。(Herberg 1963)
ラットは外側視床下部の内側前脳束に電極を取り付けられていました。
研究者は、同じ脳部位に電極を取り付けた12匹のラットを用いて、この現象を検証しました。
すると10匹が自己刺激を続ける中、定期的に陰茎から精液を放出しました。
精液の放出には、いくつかの普通でない特徴が見られました。まず、自己刺激の前中後のいずれの時点でも陰茎の勃起は見られませんでした。
精液の放出は通常、真正の射精というよりは垂れ流す形をとり、ラットは通常、中断することなくレバーを押し続けました。
明らかな射精が見られた場合、それは常に、自発的または強制的な自己刺激の終了から約5秒後に起こり、背骨が弓なり、1回か2回のけいれん的なよろめきがみられ、その後ラットは1~3分間静止したまま反応しませんでした。
しかし、通常の交尾で射精後に特徴的に見られる定型的な性器舐めは見られませんでした。
オスの交尾
内側視索前野/前視床下部
ミシガン大学の研究では、オスラットの前視床下部を電気刺激すると、性的能力が顕著に向上することが発見されました。(Vaughan and Fisher 1962)
ただし、この効果はラット30匹のうち3匹のみに生じました。
オスラットにメスラットを同伴させた後に、前視床下部への電気刺激を与えました。
すると刺激開始から数秒以内にマウントが始まり、高い頻度で継続しました。
射精後の不応期は有意に短縮され、不応期中もオスの行動はメスに向けられたままでした。また刺激中は陰茎がほぼ常に勃起していました。
刺激の終了とともに、これらの性行動は即座に終了しました。
あるラットでは、不応期が通常の平均5分以上から27秒まで短縮しました。このラットは、刺激期間の5分間で4回も射精しました。
オランダとスウェーデンの共同研究は、前視床下部の前方に位置する、内側視索前野がオスの性行動に重要であることを示しました。(Van Dis and Larsson 1971)
オスラットにメスラットを同伴させ、内側視索前野に電気刺激を与えると、オスラットの性行動が劇的に増加しました。
これらの変化は射精回数の劇的な増加、射精までの時間の短縮、射精までの挿入回数の減少、極端な不応期の短縮などで示されました。
ただし、この効果はラット11匹のうち2匹のみに生じました。
1匹は30分間のテスト中に12回射精し、もう1匹は同じ期間に17回射精しました。
射精までの時間は10秒未満に短縮されることが多く、いくつかのテストではラットは射精後すぐに再び交尾を始めました。
行動の変化は、電気刺激の開始と終了に密接に連動していました。
1匹のマウスを去勢したところ、性行動を示さなくなりました。しかし電気刺激を与えると、去勢オスは再び多数のマウントと数回の挿入を示しました。
刺激を受けている間、去勢オスの性行動は去勢前のレベルに戻りました。
オポッサムを使った実験でも、内側視索前野の電気刺激が性行動を促進することが示されています。(Roberts et al. 1967)
刺激を受けたオポッサムは、イヌのぬいぐるみに対して交尾行動を示しました。
交尾パターンの要素は、マウント、こすりつけ、噛みつき、くつろぎ、カチカチ音、勃起で、すべて自発的な性行動でもみられるものでした。
勃起を除き、これらはメスでもオスでも誘発されました。
刺激中にイヌのぬいぐるみを離すと、オポッサムはしつこくイヌにしがみつき、追いかけようとしました。
いくつかの例では、高強度の刺激で握力が非常に強くなったため、実験者はイヌを引き離すために刺激を止めざるを得ませんでした。
3匹のオスをメスと引き合わせて脳刺激を与えると、3匹ともぬいぐるみに対して現れたものと同様の行動を示し、さらに2匹はメスの肛門性器のあたりを嗅いだり舐めたりしました。
刺激がない場合、オスのうち2匹は自発的な交尾行動を示さず、3匹目は1回マウントしましたが、噛んだりこすったりはしませんでした。
内側前脳束
ピッツバーグ大学の研究グループは内側前脳束への電気刺激が性行動を促進することを示しました。(Caggiula and Hoebel 1966)
内側前脳束は外側視床下部を走行する神経繊維の束で、これに沿って欲求に関して重要な脳領域が配置されています。
まずメスのいない状態で、オスラットに外側視床下部の内側前脳束を自己刺激させると、運動性精子を含む陰茎からの分泌物が出ることがありました。
これは自己刺激のレバーを離した後の数秒後に発生し、通常は陰茎の勃起は伴いませんでした。
次にメスがいる状態で、オスラットは3分ごとにオンとオフを切り替える電気刺激を継続的に受けました。
するとラットの交尾が刺激に縛られるようになりました。つまり、刺激がオンのときはオスラットは繰り返し交尾しましたが、刺激がオフのときはめったに交尾しませんでした。

あるセッションでは、刺激が12分間オフになっている間、オスはメスから離れて休んでいました。刺激が再びオンになると、オスは10秒以内に交尾を始めました。
射精後もオスは性的に興奮したままで、刺激が終了するまで、しばしばメスの匂いを嗅いだり、つついたりし、いくつかの例では雌にマウントしました。
また食事をしていたオスは刺激を受けると食べるのをやめて、メスにマウントしました。
内側前脳束を電気刺激されたオスは、交尾する意欲が非常に高まっていました。
オスとメスと仕切りを隔てて隔離し、ケージ内の棒を押すことで仕切りを開けれるようにしました。刺激を受けたオスは仕切りを開けるために頻繁に棒を押し、メスと交尾したいという欲求を示しました。
腹側被蓋野
同研究グループは、内側前脳束に沿った脳領域の一つである、腹側被蓋野も性行動を促進することを示しました。(Eibergen and Caggiula 1973)
オスラットはメスのいる状態で、2分ごとにオンとオフを切り替える電気刺激を継続的に受けました。
すると内側前脳束の刺激と同様、オスラットの交尾が刺激に縛られ、交尾の7割~10割が刺激期間中に発生しました。特に射精は、ほぼ常に刺激がオンの期間中に発生しました。
電気刺激は、メスへの絶え間ない追跡、肛門性器の舐め、その他の接近パターンから判別される、強烈な性的興奮をもたらしました。
さらに、刺激によってオスは不応期にもメスを常に追いかけるようになりました。
この間、オスは部分的なマウントを見せました。つまり、舐めや腰振り無しに、メスの背中に前肢を乗せたり、メスの脇腹を掴んだりして、メスに交尾の姿勢をとらせました。
これが2分間の刺激期間中に30回にも及ぶことは珍しくもありませんでした。このような行動は、刺激のない期間にはほとんど発生しませんでした。
刺激を停止すると、交尾も性的興奮の兆候も即座に減少し、しばしば完全に停止しました。
アカゲサルを使った実験でも、腹側被蓋野への電気刺激で性行動が促進されることが観察されています。(Okada et al. 1991)
実験ではアカゲザルのオスとメスが、交尾することができるような状態で拘束されました。

オスは腹側被蓋野に電気刺激を受けると、すぐに対面のメスの腰に触れてつかみ 、その後、メスが尻をプレゼンティングして反応すると、オスはメスにマウンティングしました。
しかし、刺激がさらに長く続いたとしても、どのサルもマウンティングから挿入、腰振り、または射精に至ることはありませんでした。
サルは刺激が終わるとすぐにマウントをやめて、手を離しました。
メスをオスから遠ざけたり、下位のオスザルやヒトに置き換えたりすると、触りやマウンティングは誘発されませんでした。
また、メスのパートナーが魅力の低い別のメスに変更されると、触りを誘発するために必要な電流閾値が増加する傾向がありました。
中隔野
マサチューセッツ工科大学の研究では、去勢したラットの性的能力が中隔野あるいは扁桃体の刺激によって回復することが示されました。(Baum et al. 1982)
視索前野・前視床下部、尾状核・被殻、または黒質・腹側被蓋野への刺激では、このような効果はみられませんでした。
この実験では電気刺激の代わりに、男性ホルモンであるジヒドロテストステロンを中隔野あるいは扁桃体に注射するという、化学刺激が行われました。
まずオスラットを去勢しました。
続いてメスがいる状態で、オスラットの外側中隔あるいは内側扁桃体に刺激を加えると、交尾が促進されました。
これは主にマウント回数と挿入回数の増加で示され、15匹の外側中隔群と13匹の内側扁桃体群でそれぞれ統計的に有意でした。
また、射精率が大幅に回復し、射精後の不応期のいくばくかの回復もみられました。
射精率と不応期
内側扁桃体
コロラド大学の研究グループは、オスラットの内側扁桃体の刺激で、発情していないメスへの性行動を誘発できることを発見しました。(Stark et al. 1998)
刺激が無い状態では、オスは発情していないメスへほとんどマウントを行いませんでした。
しかし内側扁桃体への電気刺激をうけると、オスはメスへのマウントするようになりました。
また、メスを追跡する行動、肛門性器のあたりを嗅ぐ行動も増加しました。
これらは刺激強度が高いほど顕著でした。
同研究グループは、性経験のあるオスとないオスに分けて、内側扁桃体の性行動への影響を調べました。(Stark 2005)
性経験のないオスラットの内側扁桃体に電気刺激を加えると、オスラットへの性行動が増加しました。
刺激が無い状態ではオスはオスへほとんどマウントを行いませんでしたが、刺激をうけるとオスへマウントするようになりました。
また、オスを追跡する行動、肛門性器のあたりを嗅ぐ行動も増加しました。
一方で性経験のあるオスラットは内側扁桃体に刺激を受けても目立った変化はみられませんでした。
視床下部の背内側核
サルの視床下部の背内側核の刺激でも性行動が促進されることが示されています。(Perachio et al. 1979)
この刺激は視索前野より性行動を推進しました。
アカゲザルのオスの視索前野か視床下部の背内側核を刺激すると、メスへのマウンティングが生じ、場合によっては挿入に至りました。
マウンティングは刺激か12秒以内に発生しました。
さらに視床下部の背内側核への刺激では、挿入から射精につながりました。
自然な性行為と比較すると、刺激により、マウントの持続時間が長くなり、マウント当たりの腰振りの回数と速さが増加し、試験当たりの射精回数が増加し、射精後不応期が短くなりました。
自慰行為
延髄
アメリカ国立衛生研究所は、リスザルの脳内で射精に至る、自慰行為のような引っ掻きを誘発する脳部位を発見しました。(MacLEAN et al. 1963)
この部位の刺激により、性器の皮膚が激しく引っ掻かれ、引っ張られ、揉まれ、その後に射精が起こりました。
この部位は延髄上部のレべルにあり、内側毛帯の外側にありました。
被殻
ヤーキーズ霊長類研究所は、アカゲザルの脳内で勃起誘発部位の探索中に、自慰行為のような性器いじりを誘発する部位を発見しました。(Robinson and Mishkin 1968)
この反応は主に被殻から得られ、より少なくは前帯状皮質から得られました。
性器いじりは自慰行為的でしたが、リズミカルではありませんでした。
この活動には必ず大きな勃起が伴い、球海綿体筋と坐骨海綿体筋が収縮し、その結果、亀頭が一時的に拡張し充血しました。
しかし、射精は見られませんでした。
同研究所は続けて、勃起誘発部位の電気刺激が、無拘束のサルにどのような性的反応を生み出すのか調べました。(Perachio et al. 1969)
この実験では前視床下部と被殻が調べられました。
まずサルを椅子に拘束した状態で実験が行われました。
前視床下部を電気刺激すると、直後に動物は機敏になり、陰茎は3~5秒以内に勃起し始め、サルは実験者に向かって顎を開いて威嚇しました。
被殻の電気刺激は勃起を誘発し、勃起に伴って時折、手のリズミカルな動きがみられました。
サルは腰の柵から性器に手を伸ばすことが困難であったため、手の動きは柵より上の下半身に限定されていました。
続いてサルが拘束されていない状態で実験が行われました。
被殻の電気刺激により、手のリズミカルな動きが自慰行為に移行しました。
一方、前視床下部への電気刺激は、同じサルに大きな勃起を引き起こしましたが、自慰行為は起こしませんでした。サルは走り、叩き、顔をしかめ、刺激が消えると唇を鳴らし、耳を平らにしました。

次にオスをメスに引き合わせました。
刺激無しでは、オスは好みではないメスとは自発的には交尾しようとせんでした。
被殻を電気刺激すると、刺激開始から5~20秒以内にそのメスに対してマウントが誘発されました。
対照的に、オスが好みのメスと一緒にされると、被殻刺激は性行動を促進するようで、マウントの回数と長さが増加し、急速に射精に至りました。

一方、前視床下部の刺激では大きな勃起が得られましたが、性的能力は低下し、マウントは起こりませんでした。
メス
動物のメスでは、視床下部の腹内側核と水道周囲灰白質の電気刺激で交尾が促進されることが示されています。
また下垂体や視床下部、内側扁桃体の電気刺激で排卵や子宮収縮が生じ、さらにそれが出産や流産につながることが示されています。
メスの交尾
視床下部の腹内側核
ロックフェラー大学の研究グループは、ラットの視床下部の腹内側核へ電気刺激がメスの性受容性を高めることを発見しました。(Pfaff and Sakuma 1979)
これはメスが腰を上げて背を反らし、オスの挿入を受け入れる姿勢をとる、ロードシスと呼ばれる行動によって示されました。
まず、メスラットの卵巣を摘出した後にエストロゲンを全身投与しました。
メスは実験者による皮膚刺激か、オスラットのマウンティングに応じてロードシスを行いました。
ここで視床下部の腹内側核に電気刺激を与えると、ロードシスの頻度と強度が増加しました。
するとロードシスの頻度が刺激前には10回中4回だったのが、刺激中は10回中9回と大幅に増加しました。
オスの性行動に重要な内側視索前野の電気刺激は、逆にロードシスを抑制しました。
ロードシス
視床下部の腹内側核への刺激は、性受容性だけでなく、求交尾性 (性的交流へのメスの積極的な関与) も高めることが示されています。(RUBIN and BARFIELD 1980)
まず、メスラットの卵巣を摘出しました。
次にエストロゲンを視床下部の腹内側核に選択的に注入するという、化学刺激が行われました。
続いてプロゲステロンの全身投与し、その後オスラットに引き合わせました。
するとメスはオスのマウンティングに応じて、ロードシスを行いました。
それだけでなく、メスラットは突進、跳躍、耳を振るなどといった、誘惑する行動を示しました。
視床下部の腹内側核の近隣部位の刺激は軽度の効果を示し、そこから外れた部位の電気刺激では効果はありませんでした。
性行動
サルを使った実験でも、メスの視床下部の腹内側核の電気刺激で、求交尾性が高まることが示されています。 (Koyama et al. 1988)
これはオスに向かって自発的に尻を提示して挿入を求める姿勢をとる、プレゼンティングと呼ばれる行為によって示されました。
実験ではニホンザルまたアカゲザルが用いられ、オスメスともに拘束されました。
メスは自らプレゼンティングできる一方で、オスはメスがそうしない限り交尾はできないように配置されました。
実験配置図 (Oomura et al. 1988)
ここでメスの視床下部の腹内側核を電気刺激すると、プレゼンティングが誘発されました。
電気的に誘発されたプレゼンティングは正常なものと同一でした。すなわち、ホルダーに頭を固定されながら、メスは腰を上げて臀部をオスの方に向けました。
この姿勢は、自発的なプレゼンティングと同様、触る、マウントする、腰を振るといった一連のオスの性行動を駆り立てました。
相手のオスが外されるか、メス、無生物、または実験者に置き換えられると、高強度の刺激でもプレゼンティングは誘発されなくなりました。
刺激がない場合、メスはオスに触られると反応はしましたが、自発的なプレゼンティングはほとんど見られませんでした。
水道周囲灰白質
ロックフェラー大学の研究グループは、水道周囲灰白質への電気刺激でもメスの性受容性が増加することを発見しました。(Sakuma and Pfaff 1979)
まずメスラットの卵巣を摘出した後にエストロゲンを全身投与しました。
これらのメスは実験者による皮膚刺激か、オスラットのマウンティングに応じてロードシスを行いました。
ここで視床下部の腹内側核に電気刺激を与えると、ロードシスの頻度と強度が増加しました。
水道周囲灰白質の刺激は、視床下部の腹内側核の刺激より早くロードシスを誘発しました。
ロードシス
排卵
ほとんどの哺乳類の種では排卵は周期的に自然に発生しますが、ネコやウサギなど一部の種では排卵が交尾の刺激を受けて発生します。(Dixson 2021)
どちらの種においても、脳の電気刺激で排卵が生じることが確認されています。
いくつかの脳部位の刺激で排卵が誘発されますが、最終的に下垂体前葉において排卵を誘発する黄体形成ホルモンが放出され、血液を介して卵巣に作用する、というのが共通経路であるようです。
視床下部
ケンブリッジ大学の研究では、視床下部の電気刺激によって排卵を誘発できることが発見されました。(Harris 1937)
ウサギは発情期に交尾の刺激によって排卵する種です。
発情期のウサギの視床下部の灰白隆起を電気刺激すると、15~40時間後に排卵が誘発されました。
灰白隆起は視床下部の下方にある膨張部で、その中に弓状核があり、下垂体前葉におけるホルモン放出を調節します。
UCLAの研究では、ラットの視床下部の灰白隆起で電気刺激で排卵を誘発する部位が探索されました。(Critchlow 1958)
ラットは発情期に自然排卵する種です。
発情前期のラットの灰白隆起の領域の電気刺激すると、翌朝には排卵が確認されました。
排卵を誘発した部位は、視床下部の弓状核、腹内側核、視索上核などにありました。
電気的に誘発された排卵後の子宮、膣、卵管の状態は、通常の排卵後に観察されるものと類似していました。
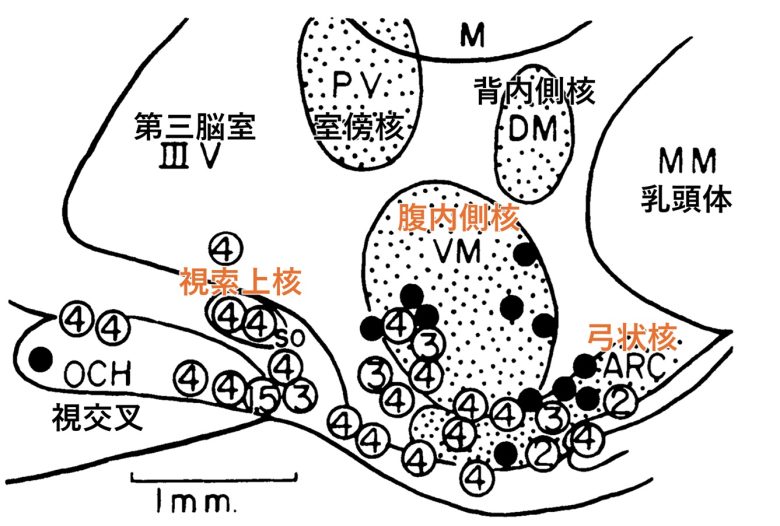
白丸:排卵成功、数字は刺激電圧。黒丸:排卵失敗。
メスヒツジを使った実験では、視床下部の腹内側核のほか、前視床下部の電気刺激で排卵が誘発できることが示されました。(PRZEKOP and DOMAŃSKI 1970)
ヒツジは発情期に自然排卵する種で、実験対象のメスは無発情期にありました。
視床下部の腹内側核、あるいは前視床下部 (視索上核と視床下部前核) を電気刺激すると、72時間以内に排卵が誘発されました。
内側視索前野
前視床下部の前方にある、内側視索前野の電気刺激でも排卵を誘発できることが示されています。(EVERETT 1965)
発情前期のラットの内側視索前野を電気刺激すると、翌朝には排卵誘発が確認されました。
対照的に、外側視索前野の電気刺激は効果がありませんでした。
また、排卵数は刺激の持続時間に比例して増加しました。
10分間の刺激では排卵が誘発されず、刺激時間が長くなるにつれ徐々に排卵数が増加し、60分間の刺激では平均10個もの排卵が誘発されました。
排卵数
電気刺激の持続時間に比例して排卵数が増加した。
内側扁桃体
日本の解剖学者の小池上春芳博士は扁桃体の電気刺激でも排卵が誘発されることを発見しました。(Koikegami et al. 1954)
発情期のウサギの扁桃体の内側核を電気刺激すると、24~48時間後に排卵が誘発されました。
この他、卵子を含んだ出血性卵胞や、新たに形成された黄体が確認されました。
博士は、扁桃体の内側核から生じた信号が視床下部の腹内側核を経由して下垂体前葉まで伝わり、そこで排卵誘発ホルモンが分泌されて卵巣に影響を与えた、と考察しました。
ラットを使った実験でも、扁桃体の内側核の電気刺激で排卵が起きる事が確認されています。(VELASCO and TALEISNIK 1969)
発情前期のラットの扁桃体の内側核に電気刺激を加えると、翌朝には排卵誘発が確認されました。
また、電気刺激から3時間後には、排卵を誘発する黄体形成ホルモンの血中濃度が大幅に上昇していたことがわかりました。
また、電気刺激による排卵は、扁桃体の内側核と視床下部の腹内側核を接続する神経繊維である、分界条の切断によって妨げられました。
したがって、信号が扁桃体-視床下部-下垂体前葉と伝わり、そこで排卵誘発ホルモンが放出されて卵巣に影響を与える、という見解を裏付ける結果となりました。
子宮収縮 (さらに出産・流産)
排卵とは異なり、脳の電気刺激によって引き起こされる子宮収縮は、2つの異なる経路を通じて発生するようです。
1つは下垂体後葉からのオキシトシン放出を通じた血液を介する経路であり、もう1つは交感神経系の活性化を通じた神経系を介する経路です。
下垂体
オハイオ州立大学の研究は、下垂体の電気刺激によって子宮収縮を誘発できることを発見しました。(Haterius and Ferguson 1938)
出産後のウサギの下垂体の漏斗部に電気刺激を加えると子宮運動が増強され、収縮の頻度と大きさが著しく増加することが観察されました。
誘発された子宮反応は50分から1時間も持続し、オキシトシン注射後に見られる反応に驚くほど類似していました。
視床下部
視床下部の電気刺激も子宮収縮を誘発できることが示されています。(Cross 1958)
授乳中のウサギの下垂体の漏斗部の他、視床下部の室傍核あるいは視索上核を電気刺激すると、子宮収縮が観察されました。
室傍核の刺激は、25~30 mUの子宮収縮剤の注射に匹敵する反応が生じることがありました。
実験はウサギの脊髄を切断した状態で行われたことから、これらの反応は下垂体後葉におけるオキシトシンの放出によるものとされました。
視索上核、室傍核にはオキシトシンを産生するニューロンが存在し、下垂体の漏斗部を通過して、下垂体後葉に終止します。
ノルウェー大学の研究者は同様の内容を確認しました。(Setekleiv 1964)
実験ではまずウサギに避妊手術を施したのち、合成エストロゲンのジエチルスチルベストロールを継続的に投与しました。
続いてウサギの視床下部の室傍核または視索上核に電気刺激を与えました。
すると、15~25秒後に最初の子宮収縮が起こり、その後、同じかやや小さい振幅で収縮が数回起こりました。
この子宮反応もオキシトシン注射後に見られるものと同様のパターンを示しました。
延髄
同研究者は延髄の電気刺激で子宮収縮を誘発できる事も示しました。(Setekleiv 1964)
ウサギの延髄の外側部を電気刺激すると、収縮とそれに続く律動的な活動の抑制からなる子宮反応が引き起こされました。
これは交感神経活性化の特徴的なパターンでした。
発生までの時間は2~4秒であり、神経系による子宮の直接的な活性化を示唆しました。
扁桃体
小池上博士は扁桃体の電気刺激で子宮収縮が起きることを示しました。(Koikegami et al. 1954)
ウサギ、イヌ、ネコの扁桃体の内側核を電気刺激すると、刺激直後か数秒後には、子宮角の運動性に顕著な変化がみられました。
すなわち、収縮の頻度と大きさが増加し、その後動きが減少または一時的に停止しました。
反応の速さから言って、これは血液ではなく神経系を介して伝達したものと考えられました。
デューク大学の研究では、妊娠した動物に扁桃体の電気刺激で子宮収縮を誘発すると、結果として出産や流産が起きえることが示されました。(Shealy and Peele 1957)
まず3匹の妊娠してないネコを対象に実験が行われました。
扁桃体の内側部および中央部への電気刺激で、子宮収縮の頻度と大きさの増加が認められました。
2匹のネコでは30秒の遅延があり、刺激終了後に2~3分間効果が持続しました。
もう1匹のネコでは、1~2分間の刺激により、20秒後に単発の、より強く持続的な収縮がみられました。4分後、収縮の頻度と振幅が増加し始め、12~15分後に最大に達し、刺激終了から約17分後に正常に戻りました。
続いて、2匹の妊娠したネコを対象に実験が行われました。
扁桃体の内側部および中央部への1分間の電気刺激を与えると、子宮収縮が始まりました。
1匹のネコは翌朝流産しました。
もう1匹は翌朝流産しなかったため、20分間の電気刺激をさらに与えたところ、翌晩に4匹の子猫を出産しました。
