快楽と欲求
脳の特定部位の電気刺激により、人間や動物に快楽と欲求を引き起こすことができます。
欲求を生じる部位は内側前脳束と呼ばれる神経線維に沿っており、中脳から大脳辺縁系にかけて広く分布しています。
快楽を生じる部位は欲求のそれに比べて少なく、「快楽中枢」あるいは「快楽ホットスポット」として知られる限られた脳領域に点在しています。
これを悪用すると、個人が本人の意思に反する行動を取るよう動機づけることも可能になると思われます。
実際、精神外科手術を拒絶していた男性が、外側扁桃体の電気刺激をうけるとその意思を翻し、手術を受けることに同意した例があります。
目次All_Pages
回路
快楽
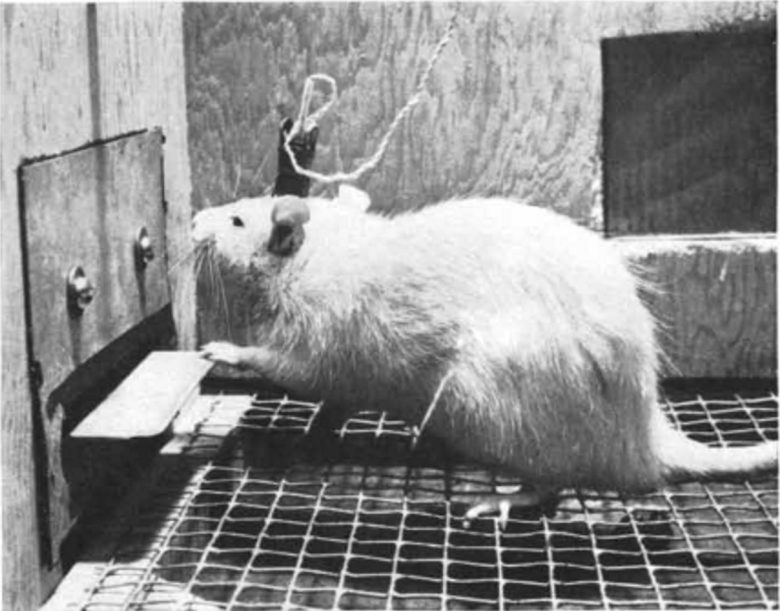
1950年代に、中隔野に電極を埋め込まれたラットが、そこへ電気刺激を加えるレバーを押し続け、強迫的に自己刺激を繰り返すことが発見されました。 (Olds and Milner 1954)
中隔野の自己刺激を繰り返した男性は、オーガズムへ向かっているかように振る舞い、「気持ちがいい」と述べました。(HEATH 1963)
中隔野にアセチルコリンとノルアドレナリンを注入された女性は、反復的なオーガズムに至りました。(HEATH 1972)
この中隔野は「快楽中枢」として知られるようになりました。
21世紀に入ると、側坐核、淡蒼球、島皮質、眼窩前頭皮質にオピオイドを注入すると、ラットの甘味への快楽的な表情が増幅されるということが発見されました。(Morales and Berridge 2020)
これらの領域は「快楽ホットスポット」ととして知られています。
このうち側坐核は欲求に関しても重要な領域で、中隔野と隣接しています。
中隔野と側坐核は相互接続しており、これが側坐核の快楽的影響をもたらす能力に寄与していると考える研究者もいます。(Zahm et al. 2012)
また、扁桃体、特に外側領域の電気刺激で人間に快楽反応が生じることが観察されています。
扁桃体の主要な出力経路の1つである腹側扁桃体遠心路は、主に扁桃体の外側領域にある中心核と基底核から始まり、視床と視床下部、そして中隔野と側坐核をつないでいます。(Kamali et al. 2015)
このつながりが扁桃体に快楽を生み出す能力を与えているものと考えられます。
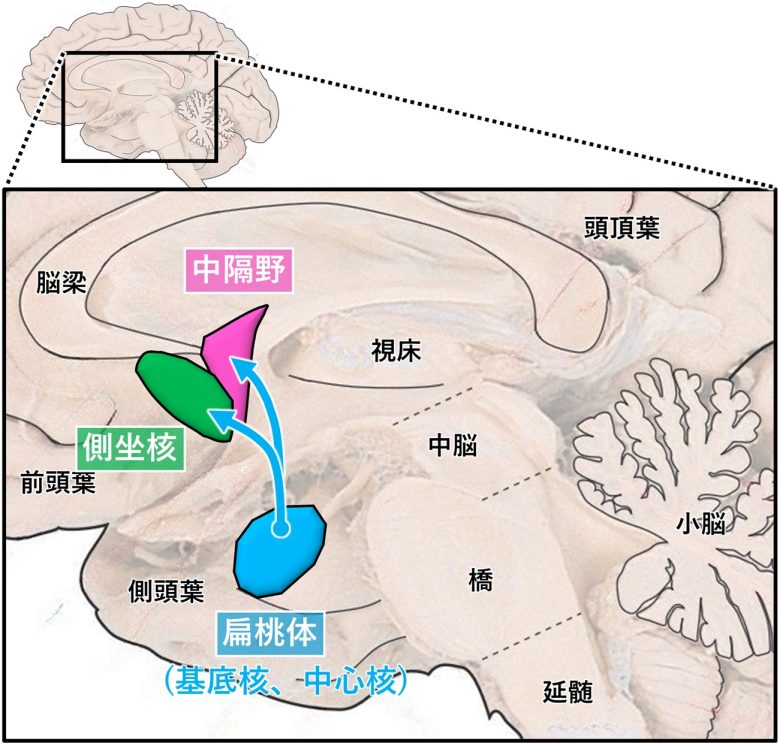
快楽に関する2つの主要な脳部位である中隔野と側坐核と、その上位の脳構造である扁桃体。
(脳画像はVanderah 2018から)
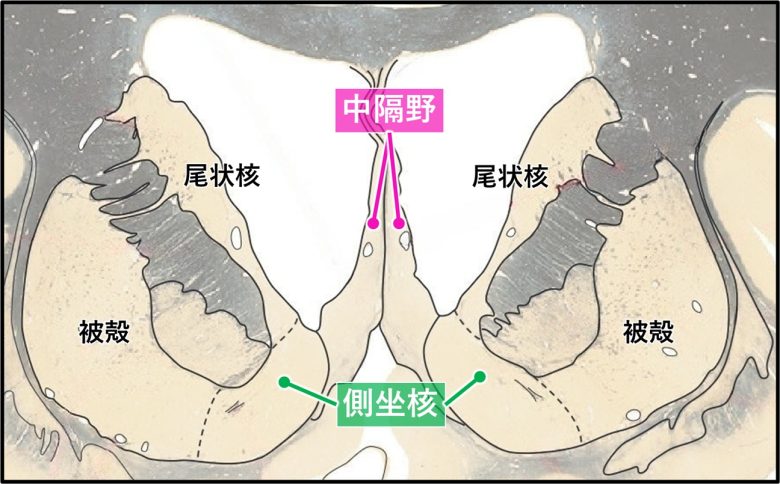
中隔野と側坐核は隣接しており、正中線上に中隔野が、その左右に側坐核が位置している。
(脳画像はVanderah 2018から)
欲求
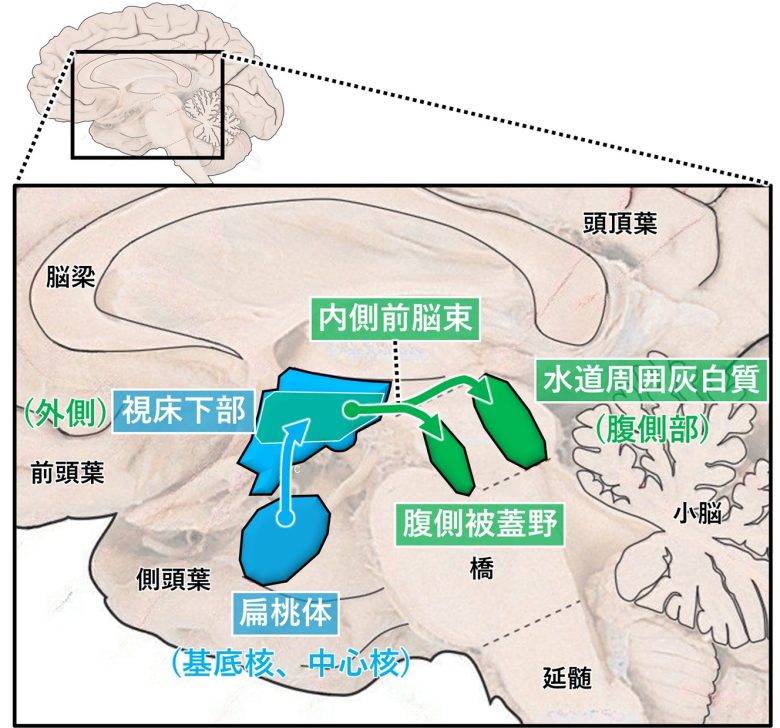
欲求に関する脳部位は内側前脳束、外側視床下部、腹側被蓋野、側坐核、扁桃体、水道周囲灰白質などがあります。
内側前脳束は外側視床下部を走行する神経繊維の束で、長さ、起源、終末が異なる50本以上の下行性および上行性の線維から構成されます。(Nieuwenhuys et al. 1982)
この神経線維に沿って、欲求に関して重要な脳構造が配置されています。(Wise 2005)
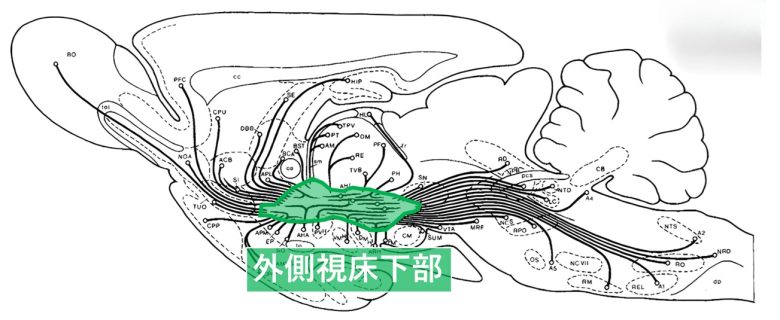
ラットの内側前脳束。外側視床下部を出入りする多数の神経線維から構成される。
(Nieuwenhuys et al. 1982を改変)
外側視床下部の内側前脳束を電気刺激すると下行性の神経繊維が活性化し、中脳の腹側被蓋野が活性化します。(Sheehan et al. 2004)
続いて腹側被蓋野にあるドーパミンニューロンが活性化し、これは逆に上行して側坐核でドーパミンを放出し、動機付けの効果をもたらします。
食物や水、性的刺激などだけでなく、乱用薬物などもこのドーパミンニューロンの活動を刺激することによって人間や動物を動機付けると広く信じられています。
扁桃体は外側視床下部の上位にある部位で、特に扁桃体の基底核と中心核は外側視床下部への強い入力があり、摂食などの動機付け行動を制御しています。(Reppucci and Petrovich 2015)
水道周囲灰白質の腹側部は、外側視床下部から広範囲に渡る入力をうけます。この領域の活動は、動物に探索、摂食、捕食行動を引き起こすのに必要十分です。(Behbehani et al. 1988, Reis et al. 2024)
以上で説明した脳領域を電気刺激すると、人間では意欲や食欲が増加し、また軽躁状態に陥ること、動物では摂食、飲水、捕食、交尾などの様々な動機付け行動が生じることが示されています。
下行性の欲求回路
(脳画像はVanderah 2018から)

上行性の欲求回路 (ドーパミンニューロン)
(脳画像はVanderah 2018から)
欲求回路と激怒・恐怖回路
これまで説明した欲求回路と激怒・恐怖回路を対比すると以下のようになります。
2ページ目で説明した、防衛的激怒と捕食攻撃の2種類の攻撃回路を汎化したものになりました。
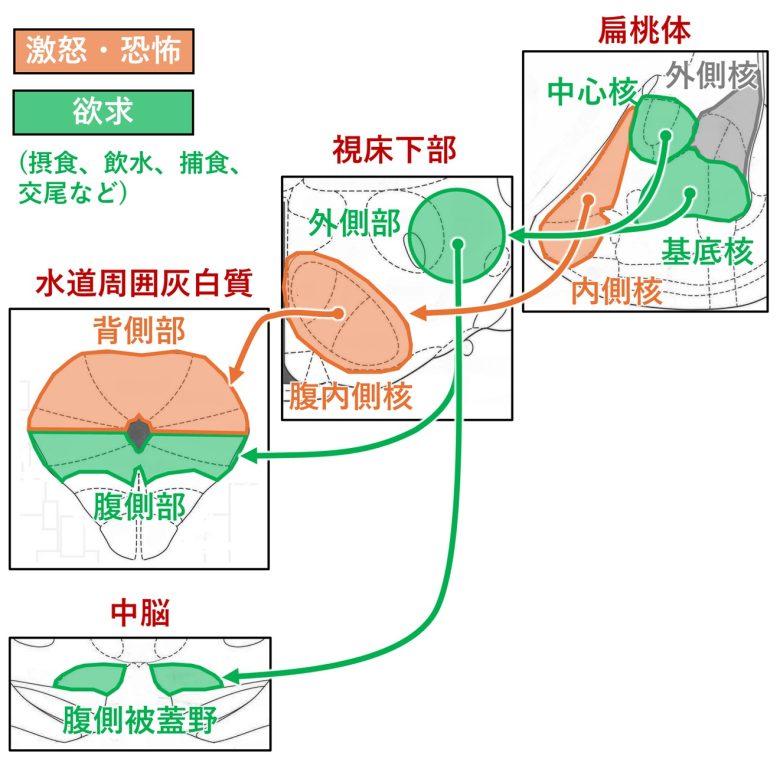
欲求回路と激怒・恐怖回路の対比
(脳地図はPaxinos and Watson 2009から)
扁桃体の2つの出力経路
扁桃体には分界条と腹側扁桃体遠心路と呼ばれる2つの主要な出力経路があります。(Isaacson 2012, Sun et al. 1991)
分界条は主に視索前野、前視床下部、そして視床下部の腹内側核へと進みます。
この神経繊維は主に扁桃体の内側核と皮質核から生じますが、基底核と中心核からもいくらか寄与があります。
腹側扁桃体遠心路は、中隔野、側坐核、視床、そして外側視床下部と接続します。(Kamali et al. 2015, Leonard and Scott 1971)
この神経繊維は主に扁桃体の基底核と中心核から生じますが、内側核と皮質核からもいくらか寄与があります。
分界条は、恐怖・激怒に関与する視床下部の腹内側核へ出力し、否定的な反応を媒介します。
腹側扁桃体遠心路は、欲求に関与する外側視床下部と、快楽と欲求に関与する中隔野・側坐核に出力し、肯定的な反応を媒介します。
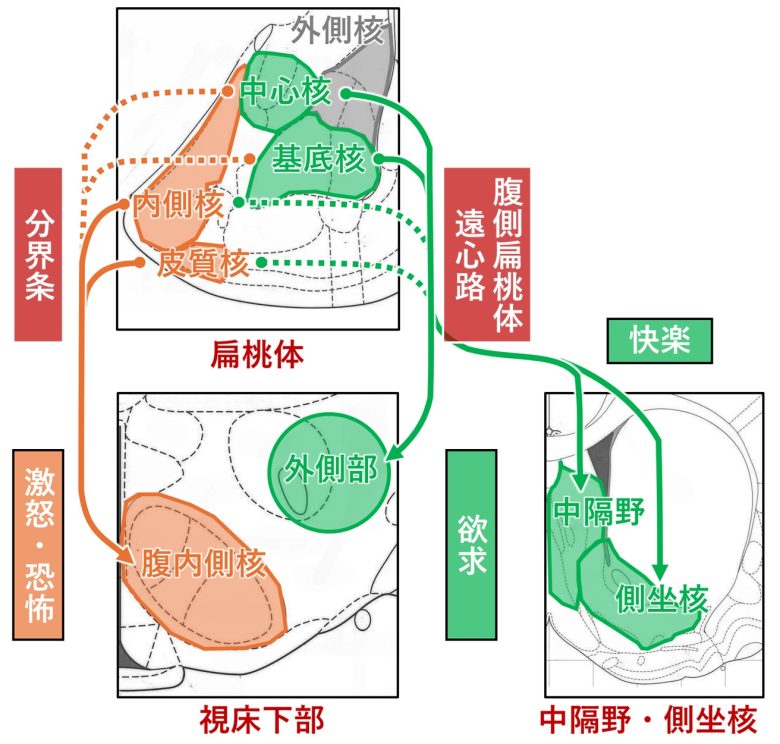
分界条と腹側扁桃体遠心路の視床下部への出力
(脳地図はPaxinos and Watson 2009から)
扁桃体を電気刺激すると、幸福感や多幸感、摂食や捕食や自己刺激といった肯定的な反応と、激怒や恐怖、攻撃や逃走という否定的な反応が、同一の領域から出現することが観察されています。
これは分界条と腹側扁桃体遠心路、どちらの神経線維が活性化されているのか、という観点から説明できるかもしれません。
少なくとも、人間の扁桃体の電気刺激で、快楽が生じた場合には腹側扁桃体遠心路が、怒りや不安感が生じた場合は分界条が活性化した実例があります。(Avecillas-Chasin et al. 2020)
人間
扁桃体、中隔野、側坐核などの電気刺激で快楽が生じることが示されています。
内側前脳束や側坐核の電気刺激で意欲が増加、また軽躁状態に陥ることが示されています。
外側視床下部の電気刺激では食欲が増加することが示されています。
快楽
扁桃体
テュレーン大学の精神科医ロバート・ヒースは、統合失調症の患者を対象にした人体実験で、いくつかの脳部位の電気刺激で快楽が生じることを発見しました。(Heath 1964)
ヒースが最も好んで刺激した脳部位は中隔野で、患者に一貫して性的な快楽反応を申告しました。(5ページ目「性行動」参照)
このほか、複数の患者が扁桃体の電気刺激で快楽が生じたことを申告しました。
35歳の男性患者 「B-12」 の脳には多数の電極が埋め込まれ、レバーを押して患者自身が脳を電気刺激できるようにしました。(Bishop et al. 1963)
各部位への電気刺激が快楽的な影響をもたらすかどうか、刺激回数によって示すよう患者に指示しました。(良くも悪くもない:3回、悪い:3回未満、良い:好きなだけ)
すると患者は中隔野、扁桃体、尾状核頭部、視床下部の自己刺激を繰り返し、これらの脳部位が「良い」ことを表明しました。
27歳の女性患者「A-8」の外側扁桃体に電極が埋め込まれ、そこへの低強度の電気刺激は快楽反応を誘発し、患者はしばしば笑いました。(HEATH et al. 1955, Heath 1996)
一方で、電流が内側扁桃体まで広がって活性化するほど電気刺激の強度を上げると、患者が抵抗できないほどの、激怒反応が誘発されました。
ハーバード大学の精神科医フランク・アービンと神経外科医バーノン・マークも同様の現象を観察しました。(Mark and Ervin 1970, Mark et al. 1972)
34歳の男性患者トーマス・R 氏は深く傷ついた気持ちと抑うつ症状を伴う、攻撃的な発作に悩まされていました。
彼の扁桃体の内側領域へ電気刺激を加えると、攻撃的な発作の前兆がみられたため、刺激は即座に停止されました。
一方、扁桃体の外側領域に電気刺激を加えると、「途方もないくつろぎと安堵感」と評される快感が得られました。
これらの快感は15~30秒の潜伏期間があり、刺激が停止した後も数分から数時間持続しました。
彼が拒絶していた精神外科手術への同意をとることさえできました。
他にも、多様な一時的な気分や感覚の変化が見られましたが、上述の快感とは対照的に、これらは電気刺激の持続時間に限定されていました。
この変化は以下のように記述されていました。
左扁桃体:「素晴らしいくつろぎ。心地よい解離。超然としており、うずき、自信に満ちている。」
右扁桃体:「心地よく、希望に満ちている。くつろいでいる。デメロール (鎮痛薬) のような快感。自信に満ちている。発作とは正反対。創造的。高揚感。非現実感。浮遊感。隔絶感。温かさ。平和。穏やか。自然で完成している。満足感。深い思考。まるで自信を持って未来のため新しい場所に行くような感じ。」
性的感覚や味覚は生じず、他の脳部位ではこのような結果は得られませんでした。
遠隔テレメトリーによってこの部位を警告無しで5回に分けて電気刺激したところ、患者がひどく落ち込んでいた日に、典型的なくつろぎと多幸感が誘発されました。
この扁桃体の外側領域の刺激が10日間中断されると、患者は怒りっぽくなり、重度の抑うつ状態になりました。
フランスのマルセイユの病院では、扁桃体の外側領域の電気刺激で生じる感情反応について、左右の扁桃体で違いがあるか調査する実験が行われました。(Lanteaume et al. 2006)
被験者は8名のてんかん患者で、主に扁桃体の外側核と基底核に電極が埋め込まれました。
左右の扁桃体の電気刺激で生じた感情反応は、自己評価式の情動尺度によって定量化されました。
すると右の扁桃体の刺激では否定的な感情のみが増加しました。
一方、左の扁桃体の刺激では喜び、幸福といった肯定的な感情も生じました。
定量化された感情反応
UCLAの研究者は、扁桃体の外側領域の電気刺激で生じる感情反応に加えて、活性化される経路も調査する実験を行いました。(Avecillas-Chasin et al. 2020)
被験者は2名のPTSDの患者で、主に扁桃体の外側核と基底核に電極が埋め込まれました。
扁桃体の基底核を電気刺激すると、幸福感、多幸感、くつろぎといった肯定的な反応が生じました。
この刺激では腹側扁桃体遠心路という、外側視床下部や中隔野、側坐核につながる経路の活性化が確認されました。
対照的に、扁桃体の外側核を電気刺激すると、怒り、不安といった否定的な反応が生じました。
この刺激で分界条という、内側視床下部につながる経路の活性化が確認されました。
また怒りは、比較的高い刺激強度と関連していました。
側坐核
スイスの精神科医のトーマス・シュレープファー博士は、側坐核への刺激で過度の多幸感が生じた症例を報告しました。(Synofzik et al. 2012)
患者は全般性不安障害と強迫性障害の複合型と診断された33歳のドイツ人男性で、外部の医療センターで側坐核への電極を埋め込まれていました。
彼は刺激装置の電池交換と刺激パラメータの調節のために博士の病院を訪れました。
パラメータ調節時に2つの興味深い現象が明らかになりました。
第一に、刺激の強さに比例して幸福感とくつろぎ感が増加しました。
刺激の強さの増加に伴い、患者はこの経験を「ハイになる」または「ヤクをやっている」感になぞらえることもありました。
患者の気分の改善は、患者の快楽的な振舞いや大きな笑顔からも明らかでした。
第二に、より強い刺激では、これらの感覚は過剰、あるいは「やりすぎ」になりました。
患者は「非現実的なほど良い」と感じ、幸福感と安らぎの感覚に「圧倒」され始めました。
彼は、その良い効果が「崩れ去り」、「不安が再発」するのではないかと恐れるようになりました。
これらの所見に基いて、中程度の刺激強度が合意され、患者は比較的幸せで、不安感も圧倒的な幸福感もない、くつろいだ状態になりました。
これらの結果に基づき、中程度の刺激強度が合意され、患者は不安感や圧倒的な幸福感を感じることなく、比較的幸せでくつろいだ状態を保つことができました。
翌日、退院の直前、患者は次の数週間の間に「もう少し幸福感を感じたい」と感じたため、再び刺激の強さを上げるよう求めました。
依存症などのリスクと倫理的な問題の検討の結果、パラメータ設定は変更されませんでした。
尾状核
イェール大学のホセ・デルガドは尾状核の電気刺激で多幸感が誘発された症例を報告しました。(Delgado et al. 1972)
被験者は35歳のてんかんの男性患者で、側頭葉に脳腫瘍が発見されていました。
尾状核頭部の電気刺激により、患者の気分は30秒以内に著しく変化しました。
刺激前、彼は控えめで、会話は制限され、病気のことを気にしていました。
刺激後、彼の自発的な会話の量は2倍以上に増加し、友好的な表情や多幸感に満ちた行動が見られ、最終的には冗談を言ったり、右手で軽く叩きながら陽気な「カンテ・ホンド」風に大声で歌ったりするようになりました。
多幸感は10分ほど続き、患者は徐々にいつもの控えめな態度に戻りました。
これは3回の異なるセッションで再現されました。
軽躁状態
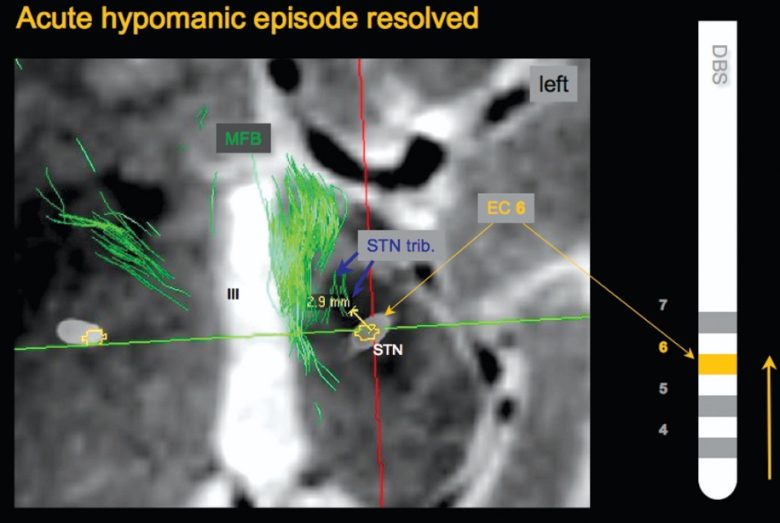
内側前脳束 (視床下核付近)
パーキンソン病において、視床下核への電気刺激は一般的な治療法ですが、その副作用として軽躁状態・躁病が生じることがあります。
具体的な症状は、異常な気分の高揚と活動の増加、怒りっぽさ、性欲の増加、妄想的な考えなどです。
この副作用は、視床下核から伸びる内側前脳束の活性化が原因であることが実証されています。
スペインの49歳の男性患者は、視床下核への電気刺激を開始してから2時間後には多幸感を表明しました。(Kulisevsky et al. 2002)
48時間後には病的な多弁、過活動、および性欲の増大も示しました。
患者は、なぜもっと早く手術を受けられかったのかと繰り返し尋ねました。
患者は、「こんなに気分がいいのは初めて」なので、すぐに退院することを要求し、商談を終えるために飛行機に乗りたいと主張しました。
躁病的な症状は一日中持続しました。彼はまた誇大妄想を抱くようになりましたが、幻覚はありませんでした。
数日後に刺激パラメータが変更されると、多動性と気分障害は部分的に改善し、患者は気分安定剤なしで退院しました。
そして退院後、軽躁病は数日で解消しました。
ドイツの65歳の女性患者は、視床下核への電気刺激を開始してから著しい気分変化を示しました。(Herzog et al. 2003)
運動機能の改善と並行して、彼女の気分は患者にとって異常なほどに高揚しました。
彼女は過度に饒舌で、話している最中に中断することは不可能でした。その後3週間にわたって、軽躁状態が精神病の症状を伴う、躁病の症状に徐々に変化しました。
患者の気分は多幸感にあふれ、話すスピードは速くなり、思考は散漫で、突飛な考えを思いつき、話が脱線し、集中力が低下しました。
彼女は社会的な抑制を失い、2人の神経科医に対して恋に落ち、人と抱き合ったりキスをしようとしました。
彼女は多動で落ち着きがなく、許可なく何度もクリニックを出たり、無節操に服を買ったりしました。乱雑な行動から部屋を散らかし、隣人のベッドを占拠していました。
彼女が経済的に破綻するのを防ぐため、家族はクレジットカードを取り上げたいと考えていました。彼女の判断力は低下しており、自身の障害に対する自覚が薄く、ストレス耐性も低いものでした。
また彼女は疑い深く、ピリピリして敵対的になり、息子が彼女に陰謀を企てているという妄想を抱き、力ずくで脅してお金を取ろうとしたと言いました。
薬物治療を併用することで、最終的にこの感情障害は解消しました。
スウェーデンの59歳の男性患者は、手術前は患者は非常に静かで無口な人物で、精神疾患の病歴はありませんでした。(Kim et al. 2012)
視床下核の電気刺激の開始後、運動障害は急速かつ顕著に改善しました。
しかし2日後には、目を見張るような気分の変化が起こりました。
彼は気分が高揚しておしゃべりになり、過度に希望的になりました。
彼は脳神経外科の病棟の他の患者と踊り回ったり歌ったりし、さらに性的関心が高まりました。
ブリティッシュコロンビア大学の研究グループは、視床下核への電気刺激で躁病が生じた3つの症例を報告しました。(Mandat et al. 2006, Coenen et al. 2009)
一人目は72歳の男性患者で、電気刺激により運動機能が著しく改善しました。
患者は副作用を申告せず、新しい運動能力のおかげで幸福感が増したと述べました。
しかし、彼は足の変形のため運転できないにもかかわらず、新車を購入しました。
また、患者は売春婦を老人ホームに呼び寄せました。この行動は、彼の性格にも施設の規則にもまったく合わないものでした。
刺激パラメータの変更によって、運動機能の改善を維持したまま軽躁状態を解消することができました。
二人目は45歳の男性患者で、こちらも電気刺激での運動機能の改善は顕著でした。
彼は副作用は報告せず、より目が覚めて注意力が増したと述べました。
しかし、患者は混雑した道路の真ん中で駐車中の車に衝突しました。彼はなぜそんなことをしたのか説明できませんでした。
事件の最中、自分が「限界を超えて無敵」だと感じていたことを覚えており、車に衝突している時、怒りは感じていませんでした。
その後に控えている裁判に対して何のストレスも感じませんでした。
刺激パラメータを変更すると、無敵感は3~4日で消え、裁判手続きに対して適度に不安を感じるようになりました。
その後、軽躁病の行動はみられなくなりました。
三人目は66歳の男性患者で、視床下核への電気刺激を開始すると、患者の歩行能力が向上しました。
しかし10日後から、患者に異常な行動が始まりました。
患者は妻の服を着るようになり、毎日性交を要求し、特に無謀な運転など、リスクを冒す行動が増加しました。
そして患者は入院しました。彼は動揺しているように見え、自分の行動がおかしいことを理解していましたが、「何かが私を駆り立てている」と説明しました。
刺激がオフになると、彼の衝動はほぼ瞬時に止まりました。
1年前、プラミペキソールというドーパミン作動性の薬を服用していた時も、似たような、しかし軽度の性的なフェチ行為が一時的に起きていました。
より浅い電極接点を使用することで、彼の運動症状は改善し、軽躁病の行動は消失しました。
同研究グループは、視床下核に電気刺激を施したとき、なぜ患者が軽躁病を発症することがあるのか、原因を調査しました。(Coenen et al. 2009)
患者の脳の神経線維を可視化すると、軽躁病を発症した患者は電極接点が内側前脳束に直接接触していることが分かりました。
一方で軽躁病を発症しなかった患者は、電極接点が内側前脳束から離れていました。
電極が接触していた内側前脳束は、よく知られた腹側被蓋野から出る本流ではなく、視床下核から出る支流でした。
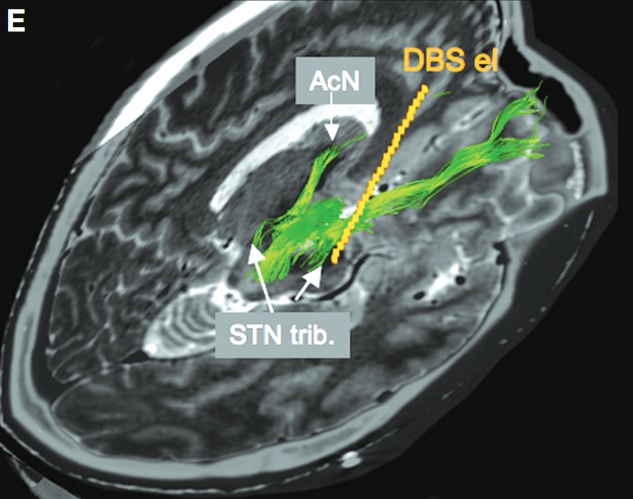
軽躁病を発症した患者の電極接点を、内側前脳束に接触しないように1段階上に変更すると、軽躁病は解消し、運動症状はさらに改善しました。

側坐核
強迫性障害の治療として側坐核の電気刺激が実験的に行われていますが、副作用として患者に軽躁状態が生じることが観察されています。
ブラウン医科大学とクリーブランド・クリニックの共同研究では、側坐核のすぐ上にある内包の電気刺激で、10人のうち5人に軽躁状態が生じました。(Greenberg et al. 2006)
刺激から数秒以内に患者の気分は高揚し、エネルギー、発話量、社会交流における自発性が著しく増加しました。
ドイツのケルン大学の研究では、側坐核の電気刺激で、10人中2人に軽躁状態が生じました。(Huff et al. 2010)
また1人の患者は、集中力の低下と記憶力の低下を申告しました。
アムステルダム大学の研究では、側坐核の電気刺激で、16人中8人に軽躁状態が生じました。(Denys et al. 2010)
また7人の患者から性欲の増加が報告されましたが、不快感はありませんでした。
意欲の増加
側坐核
うつ病の症状の一つは意欲 (やる気) の低下ですが、側坐核の電気刺激を受けた患者の意欲が増加することが示されています。
スイスの精神科医のトーマス・シュレープファー博士は、うつ病の実験的治療法として、3人の患者に側坐核への電気刺激を施しました。(Schlaepfer et al. 2007)
すると2人の患者で、即時的で急激な意欲の増加がみられました。
1人目の患者は、刺激の直後に自発的に、自分が有名なケルン大聖堂を訪れたことがないため、近い将来に訪れる予定であると述べました。
2人目の患者は、刺激の直後に自発的に、ボウリングを再開したいと述べました。これは12年前、うつ病が発症する前に好きだったことでした。
彼女は「これはかなり楽しいだろう」と述べました。
これらの患者は長い抑うつ状態の間、意欲を顕著に欠いていたため、この効果は目を見張るものがありました。
うつ病の症状は電気刺激によって3人の患者全員で改善しました。
続けて博士は、側坐核の電気刺激による長期的な治療効果を検証しました。(Bewernick et al. 2012)
11人の患者のうち5人に症状の改善がみられ、彼らは一貫して意欲の増加を示しました。
患者たちはパートタイムで働き始め、個人的な用事を片付け、新しい趣味を再開し、ある患者は子供が欲しいという願望を抱くようになりました。
2人の患者は、気分の高揚と睡眠時間の減少という、一時的な軽躁病の症状を呈しました。
内側前脳束
またシュレープファー博士は、うつ病の実験的治療として、内側前脳束への電気刺激を7人の患者に施しました。(Schlaepfer et al. 2013)
術中のテスト刺激では、すべての患者が一貫して意欲の増加と気分の改善を示しました。
意欲の向上は、定位反応、アイコンタクトの開始、心理学者との会話への参加などによって示されました。
継続的な電気刺激を開始した直後、6人が気分、不安、意欲の急激な変化を報告し、数か月後には4人がうつ病が寛解したと診断されました。
食欲の増加
視床下部
デンマークのコペンハーゲンの病院では、肥満の実験的治療として、外側視床下部の電気凝固術が適用されました。(Quaade et al. 1974)
被験者は女性4名と男性1名で、全員が過食症で体重が118 kgから180 kgにおよぶ著しい肥満でした。
この凝固術に先立って、外側視床下部への電気刺激の実験が行われました。
すると5人のうち3人が説得力ある空腹反応を示しました。
例えば以下のような発言がみられました。
「とても空腹なので、フライドポテトをお供にフライドチキンを丸ごと1羽食べられます。」
「とても空腹なので、お腹全体が真空のように感じます。」
その他、電極軌道の下部からは恐怖が得られ、上部からは多幸感に近い強烈な快感が得られました。
続いて、空腹反応が誘発された部位の凝固が行われました。
術後1週間で、患者たちは初めてあまり空腹を感じなくなり、特に食事の初めから満腹感を覚えたと述べました。
患者の体重はわずかに一時的に減少しましたが、有意な影響はありませんでした。
ある患者は、2回目の凝固後、自発性と積極性がわずかに低下しました。この状態は2~3か月続きましたが、その後、通常の精神状態に戻りました。
動物
中隔野、扁桃体、側坐核、外側視床下部、腹側被蓋野、水道周囲灰白質など、内側前脳束に沿った脳領域を動物は強迫的に自己刺激します。
このような自己刺激を繰り返した動物が薬物依存症のような症状を呈することも観察されています。
またこれらの脳領域の電気刺激によって、摂食、飲水、捕食など、様々な動機付け行動を動物にとらせることができます。
このような効果によって、例えば満腹の動物に摂食を強制させて肥満化させることもできます。
自己刺激
中隔野
1954年、アメリカの心理学者ジェームズ・オールズ博士は、中隔野に電極を埋め込まれたラットが、そこへ電気刺激を加えるレバーを押し続け、強迫的に自己刺激を繰り返すことを発見しました。 (Olds and Milner 1954)
中隔野を自己刺激している動物は、外見上、非常に満足のいく快楽を味わっているように見えました。彼らは、その経験が著しく強烈であるかのように、定期的に震えました。(Panksepp 1998)
この効果を利用して、ラットを特定の場所に誘導することもできました。(Olds 1956)
たとえば、ラットがケージのある角に行くたびに中隔野に電気刺激を与えることで、博士はラットをその角に留まらせることができました。
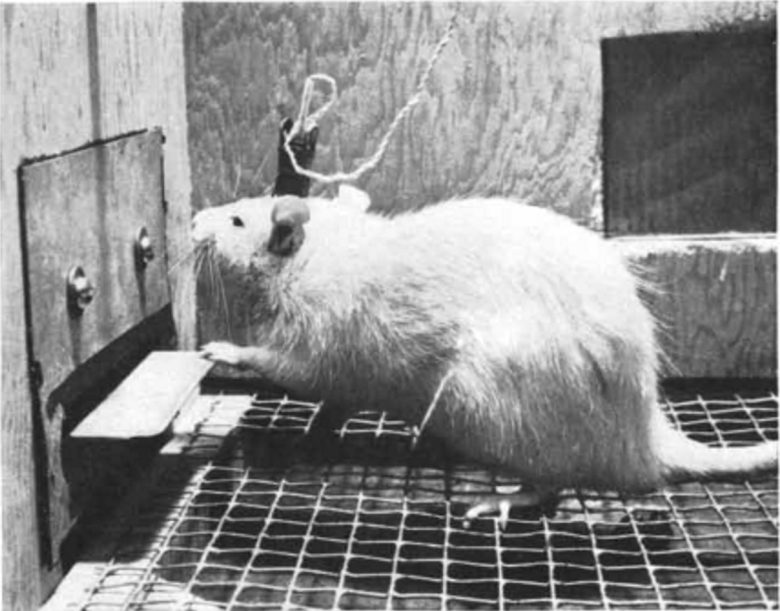
この中隔野は「快楽中枢」と呼ばれるようになりました。
この発見はラットで何度も再現され、金魚、モルモット、バンドウイルカ、ネコ、イヌ、ヤギ、サルなど他の種も同様に脳の自己刺激をすることが示されました。(Bishop et al. 1963)

椅子に拘束されながら、左手でレバーを押して脳への自己刺激を続けるサル。(Brady 1961)
驚くべきことに、脳の自己刺激の実験は人間でも行われました。
中隔野を自己刺激した被験者は、一貫して性的快楽あるいは性的欲求を体験したことを報告しました。強い快感から痙攣を起こすことすらありました。(5ページ目「性行動」参照)
内側前脳束
中隔野以外の脳領域でも、ラットが自己刺激をする領域がないか、オールズ博士は探しました。(Olds 1956, Olds and Olds 1963)
それらは、視床下部、扁桃体、側坐核、前帯状皮質、海馬、腹側被蓋野など、幅広い領域に見つかりました。
なかでもラットが最も激しく自己刺激したのは、外側視床下部から腹側被蓋野へと至る、内側前脳束に沿った間脳・中脳の領域でした。
この領域の電気刺激は、食べ物などの通常の満足感を与えるものよりも、動物にとってはるかに大きな満足感を与えるようでした。(Olds 1956)
たとえば、空腹のラットは、食物に近づくよりも自己刺激装置に近づく方がより速く走りました。実際、空腹のラットは、電気刺激による快楽を優先して、食べ物を無視することがよくありました。
中には24時間連続で1時間に2,000回以上も脳を刺激するラットもいました。
満腹ラットが脳刺激の報酬を求めて走る場合、24時間空腹のラットが餌の報酬を求めて走る場合より、報酬に向かう途中で肢に受ける電気ショックにはるかによく耐えました。(Olds 1958)
外側視床下部
コーネル大学の研究者は、中隔野と外側視床下部から得られる、ラットの自己刺激の性質を比較しました。(Spies 1965)
実験ではラットの中隔野あるいは外側視床下部のいずれかに電極を設置し、彼らが自己刺激か食べ物のどちからを好むか調査しました。
ケージの中に2つのレバーが設置されました。片方は脳の自己刺激用で、もう片方は餌の供給用でした。
ラットは1時間のセッション中に、自己刺激と食事のどちらも行うことができました。
すると外側視床下部ラットは、空腹にも関わらず自己刺激を選択し続け、自ら飢餓状態に陥ってしまいました。
一方で、中隔ラットは自己刺激を行う一方で、餌も食べて体重を維持しました。
また、外側視床下部ラットのうち、電極接点が内側前脳束に接触していないものは中隔ラットと同じ行動パターンを見せ、飢餓状態には陥りませんでした。
体重
外側視床下部+内側前脳束ラットは、空腹にも関わらず自己刺激を選択し続け、自ら飢餓状態に陥った。一方中隔ラットは自己刺激を行う一方で、餌も食べて体重を維持した。
自己刺激と摂食
外側視床下部+内側前脳束ラットは、空腹にも関わらず激しく自己刺激を行った。一方中隔ラットは自己刺激を行う一方で、餌も食べた。
腹側被蓋野
フランスのボルドー大学の研究者は、腹側被蓋野と外側視床下部から得られる、ラットの自己刺激の性質を比較しました。(Miliaressis and Cardo 1973)
自己刺激のレバーと、餌ペレットのレバーをそれぞれケージに設置しました。ラットは30分間のセッション中に、自己刺激と食事のどちらも行うことができました。
実験はラットが満腹から24, 48, 72時間の絶食という、様々な絶食度で行われました。
腹側被蓋野と外側視床下部、どちらに電極を装着したラットも、空腹にも関わらず激しく自己刺激しました。
ただ絶食期間が長くなるにつれて、摂食量は多くなりました。
腹側被蓋野ラットの方がやや多く自己刺激し、外側視床下部ラットの方がやや多く摂食するという、全般的な傾向がみられました。
絶食度ごとの自己刺激と摂食
絶食時にみられた特徴的な傾向として、腹側被蓋野ラットが次第に摂食を放棄して、自己刺激に移行したという点です。
一方で外側視床下部ラットは、自己刺激を次第に放棄して、摂食に移行したという逆の傾向がみられました。
絶食時の自己刺激と摂食
水道周囲灰白質
UCLAの研究者は、水道周囲灰白質の腹側部から、一貫した強力な自己刺激が得られることを発見しました。 (Liebman et al. 1973)
ラットの自己刺激の速度は1時間に6000回以上と非常に高く、しばしば激しい嗅ぎ行動が伴いました。
一方で、水道週灰白質の背側部をラットは全く自己刺激せず、行動の兆候から判断すると刺激が嫌悪的であることは明らかでした。
実際の所、水道周囲灰白質の背側部は激怒および恐怖反応を誘発する領域として知られています。(2ページ目「激怒」、3ページ目「恐怖」参照)
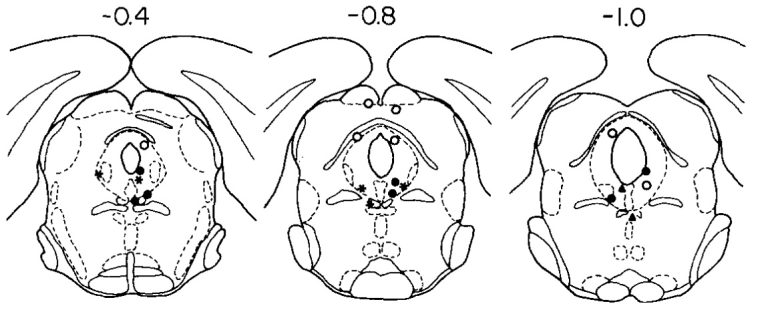
黒丸:高頻度、黒星:中程度、白丸:無-低頻度。
側坐核
シドニー大学の研究者は、側坐核と外側視床下部から得られる、ラットの自己刺激の性質を比較しました。(Jenkins et al. 1983)
外側視床下部ラットにおける活発な自己刺激の獲得は非常に速く、わずか3日しかかかりませんでした。
一方、側坐核ラットが同等の活発な自己刺激を獲得するまでには20日以上かかりました。
しかしその後、側坐核ラットはより強迫的な行動を示し、刺激に応じた嗅ぎ、立ち上がり、探索行動の欠如が顕著になりました。
また、自己刺激を1日剥奪すると、側坐核ラットは刺激装置に駆け寄り、刺激開始までの時間が大幅に短縮しました。
さらにラットは試験の3週間で徐々にけいれん発作を起こし、その後、発作はより頻繁で重度になりました。
モルヒネ離脱症候群の特徴とされる、ウェットドッグシェイクと呼ばれる運動もみられました。
これらの現象は側坐核におけるオピオイド系の変化によるものと考えられました。
つまり側坐核の自己刺激を繰り返したラットは、薬物依存症に似た症状を呈しました。
ドーパミンニューロン
スイスのジュネーブ大学の研究では、ドーパミンニューロンを継続的に活性化することで、ラットに薬物依存症でみられるような行動および細胞の異常が生じることが発見されました。(Pascoli et al. 2015)
まずラットの腹側被蓋野のドーパミンニューロンを直接レーザー光で活性化するレバーをケージに設置しました。
続いてラットがケージに入れられ、ラットはレバーを押して自身のドーパミンニューロンを自己刺激することをすぐに学習しました。
刺激のたびに肢への電気ショックを与えても、ラットは自己刺激を止めませんでした。
この肢への電気ショックは、少なくとも甘い水をもらえるレバー押しを完全に抑えるほどには強力なものでした。
ラットは12日間連続で自己刺激装置の使用が許可され、それから30日間の「禁欲」の後、再びラットを同じ装置に曝しました。
するとラットは自己刺激を追い求め、スイッチが切れた装置のレバーを押し続けるという、離脱症状のような行動をみせました。
この離脱ラットの側坐核におけるニューロンの変化は、コカイン自己投与からの離脱後に観察されるものと区別がつきませんでした。
扁桃体
当時、激怒や恐怖といった負の感情への関与でよく知られていた扁桃体を、動物が自己刺激をすることに驚いたオールズ博士は、これについて詳しく調査しました。(Wurtz and Olds 1963)
ラットの扁桃体の幅広い領域に電極が埋め込まれ、自己刺激と逃避のいずれかの反応を生み出すか調べました。
すると自己刺激はほとんどが中心核と内側核から生じ、逃避はほとんどが外側核と基底核から生じました。
しかし反応は厳密には局所的でなく、自己刺激と逃避の分布は重なり合っており、内側核から逃避が生じることもあり、また基底核から自己刺激が生じることもありました。
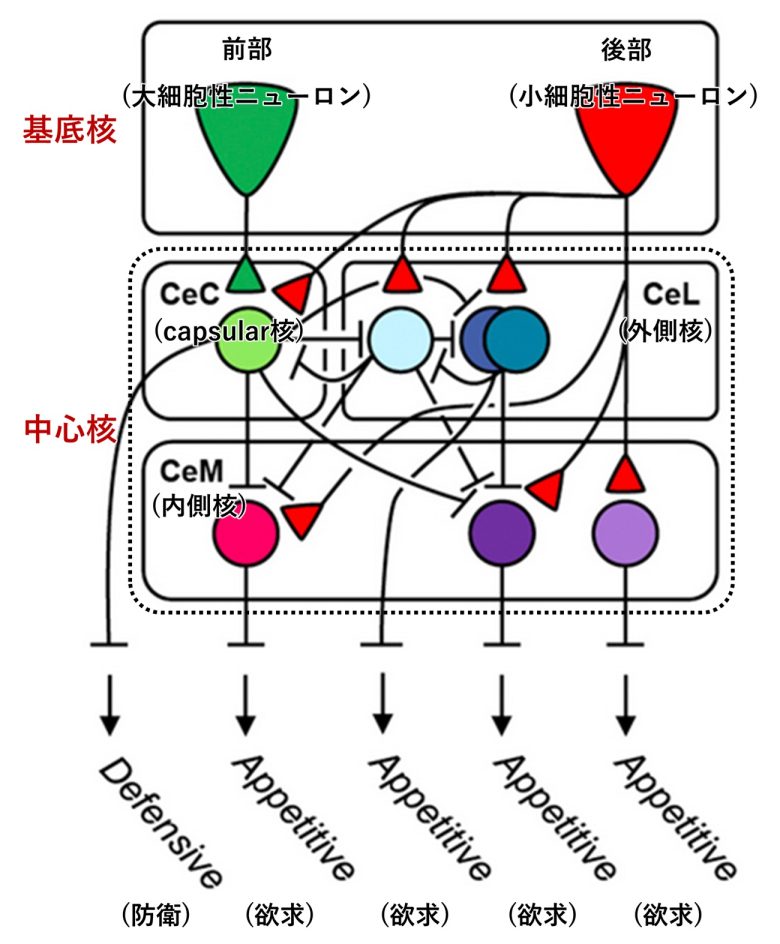
正確なレーザー刺激を用いた最近の研究は、領域のさらなる細分化により、扁桃体から得られる相反する行動を地形的に異なる領域にマッピングできることを示しました。(Kim et al. 2016, Kim et al. 2017)
実験では扁桃体の基底核と中心核をさらに細分化し、ラットの自己刺激と凍り付き反応がこれらの小領域から個別に誘発できることを示しました。
基底核前部の大細胞性ニューロンからは凍り付き、基底核後部の小細胞性ニューロンからは自己刺激が得られました。
中心核のうち、capsular核からは凍り付き、内側核と外側核からは自己刺激が得られました。
摂食
摂食行動の誘発は主に、「空腹中枢」とも呼称される、外側視床下部で確認されています。
扁桃体、内側前脳束、中隔野、水道周囲灰白質、側坐核などでもみられ、動物が自己刺激する脳領域と重なりがあります。
外側視床下部
スイスのチューリッヒ大学のヴァルター・ヘス博士はネコの外側視床下部を電気刺激すると、過食が生じることを発見しました。(Hess 1958)
それまでネコは牛乳も肉も食べていませんでしたが、刺激を受けると貪欲に食べたり飲んだりしました。
実際のところ、ネコは鉗子、鍵、棒など、食物としては不適当な物体を口に入れたり、かじったりすることもありました。
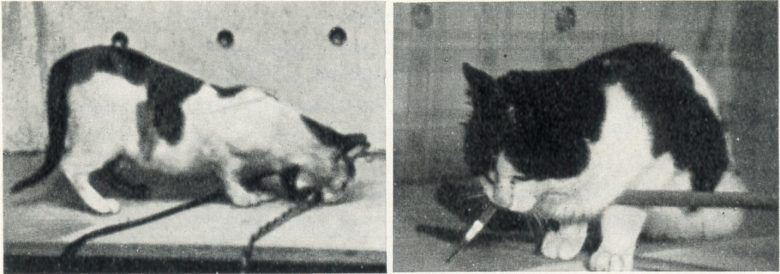
日本の研究者もネコの外側視床下部の電気刺激で過食が生じることを示しました。(UMEMOTO and KIDO 1967)
ネコはケージ内に設置された容器から餌 (魚と米) をいつでも食べることができ、十分に食事を与えられた状態でした。
外側視床下部の電気刺激を与えると、ネコは匂いを嗅ぎながら食器に近づき、魚を元気に食べました。
摂食は刺激を止めるとすぐに止まりました。このとき、ネコは数秒間、固まった姿勢を示しました。
刺激の強度が増すと、反応はより激しくなり、ネコは食べられるものと食べられないものを区別できなくなり、食器を噛んでしましました。
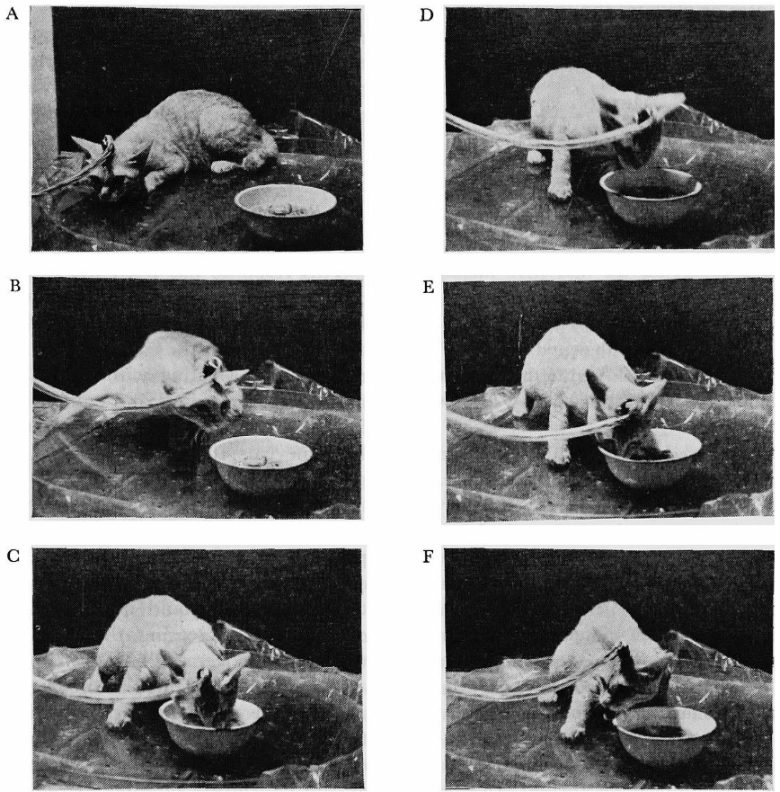
A:刺激前、B-E:刺激中、F:刺激終了直後。
イェール大学のニール・ミラー博士は過食を推し進め、外側視床下部の長期的な電気刺激によってラットを肥満化させることに成功しました。(Steinbaum and Miller 1965)
実験では1日2時間、31日間、継続的にラットの外側視床下部に電気刺激を加えました。
すると8日目からラットは刺激時間帯に過食するようになり、最終的に肥満になりました。
実験終了時には、刺激を受けたラットは1日あたり、正常な対照ラットが食べる餌の約2.5倍を摂取していました。
摂食量と体重
ラットは食物で膨満してしまい、刺激の合間では完全に活動せず、ほとんどの時間を仰向けになって、手足を空中に上げて過ごしていることが観察されました。
これらのラットの胃は異常に膨張しており、一部のラットには出血もみられました。
刺激が停止するとラットの摂食量は通常時の10%と著しく減少し、過食を代償しようとしました。
しかしラットはその後も、脳刺激によって代償できないほどの大量の過食を強制され、自由意志による摂食行動が止められ、肥満になってしまいました。
オポッサムでも、外側視床下部の電気刺激によって摂食が生じる事が示されています。(Roberts et al. 1967)
誘発された摂食行動は通常の食事でみられるものと同様でした。
食物は切歯と犬歯でつかまれ、口の奥に移され、咀嚼され、通常は飲み込まれました。
食物が口から落ち始めたら、前足を使って食物を口の中に押し戻すのが一般的でしたが、咀嚼中に食物の大部分が口から落ちてしまう場合もいくつかありました。
扁桃体
扁桃体の刺激では摂食を誘発するだけでなく、摂食を促進することも示されています。(Montgomery and Singer 1975)
この実験では電気刺激の代わりに、神経伝達物質であるノルアドレナリンをラットの扁桃体の皮質核と外側視床下部に注入するという化学刺激が行われました。
まず満腹のラットにおいて、外側視床下部の刺激によって摂食量が増加することを確認しました。
その後、さらに扁桃体の皮質核も刺激すると、摂食量の増加が強化されることがわかりました。
扁桃体の刺激だけでも摂食量の増加が確認されましたが、外側視床下部に比べると程度は小さいものでした。
摂食量
扁桃体の刺激は視床下部外側の刺激で誘発される摂食を促進した。
腹側被蓋野
腹側被蓋野の電気刺激でもラットに摂食が誘発されることが示されています。(Waldbillig 1975)
反応は視床下部から誘発されたものと類似していました。
摂食行動は、前歯で食物をつかみ、それを前肢に移し、小さな一口を齧り、咀嚼し、飲み込むという一連の動作で構成されていました。
反応が不適切な対象に向けられることはありませんでした。
中隔野
中隔野の電気刺激でもラットに摂食が誘発されることが示されています。(Altman and Wishart 1971)
刺激を受けたラットは最初に震え、続いてかなり激しく毛づくろいし、最後に食物があればそれを摂取するという一連の行動を行いました。
中隔野の刺激による摂食行動は、外側視床下部の刺激や、自然の空腹による摂食行動ほど強くないようでした。
側坐核
側坐核の電気刺激でもラットに摂食が誘発されることが示されています。(van der Plasse et al. 2012)
側坐核はシェルとコアの2つの構造に分けられますが、摂食が誘発されたのは、側坐核シェルの内側部への電気刺激のみでした。
また電気刺激の強度が高くなるほど、ラットの摂食量は増加しました。
摂食量
内側前脳束
ヤーキーズ霊長類研究所では、アカゲザルの脳に多数の電極を埋め込み、摂食を誘発する領域を調査する実験が行われました。(Robinson and Mishkin 1968)
最も低い刺激強度で反応が得られたのは、外側視床下部を中心とした、内側視索前野から腹側被蓋野にかけての、内側前脳束への刺激でした。
典型的な反応では、サルは電気刺激の開始から2~5秒後に、口の近くに置かれた食物を摂取しました。
激しい反応では、食べ物はすぐに頬袋に運ばれ、その後、頬袋と口の両方がいっぱいになり、サルがそれ以上受け取ることが物理的に不可能になるまで、さらに多くの食べ物が受け入れられました。
それほど強くない反応では、食物は口の中にのみ保持されました。
反応の強さに関係なく、受け取った食物は刺激中に噛まれて飲み込まれることはほとんどなく、刺激の停止後3~15秒以内に定期的に頬袋と口から排出されました。
刺激を繰り返すたびにこれらの行動は再現されました。
また、食物以外の物体の摂取が起きる事もありました。この行為は食物の摂取に非常に似ていました。
激しい反応では、口と頬袋がコルクやその他の物体でいっぱいになることもあり、これらの物体はいずれも飲み込まれませんでした。
捕食
ここでは摂食と捕食が同一の脳領域の電気刺激から生じた例を紹介します。
その他の捕食攻撃の実験については、2ページ目「激怒」をご覧ください。
外側視床下部
イリノイ州の精神病院の実験では、ネコの外側視床下部の電気刺激で、摂食だけでなく、ラットへの捕食攻撃が生じる事が示されました。(Hutchinson and Renfrew 1966)
まず実験開始の1時間前に、ネコに1日分の食事をさせました。
続いてネコをラットと一緒にケージに入れました。自発的にはネコはラットを攻撃しませんでした。
ここでネコの外側視床下部を電気刺激すると、ネコはラットに対する捕食攻撃を開始しました。
ネコはケージの床近くまでかがみ、ラットに忍び寄り、ラットの頭または首を顎でつかみ、片方の足でラットを床に釘付けにしました。
攻撃は無音で、瞳孔の拡張、尾に限定した毛の逆立ち、呼吸の増加を伴いました。刺激が終了すると、攻撃はすぐに止まりました。
続いてケージにラットの代わりに馬肉の皿を置きました。
ここでネコの外側視床下部を電気刺激すると、ネコは肉を食べ始めました。
ネコは刺激から15 -20秒後に皿に近づき、刺激期間中ずっと食べ続けました。
ラットと馬肉、どちらもある状態で刺激を受けると、ネコの捕食攻撃と摂食のどちらも誘発されました。
ラットを捕食するか、馬肉を食べるかは、どの目標に最初に接触したかによって決まりました。
ラットがネコにより近くに配置されていた場合、ラットがネコから攻撃されました。
逆に皿がより近くに置かれていた場合、皿の馬肉が食べられました。
水道周囲灰白質
UCLAの研究者は、マウスの水道周囲灰白質の腹側部の刺激により、探索、摂食、捕食が生じることを示しました。(Reis et al. 2024)
ケージに何も存在しないときは探索行動が、クルミの存在下では摂食が、コオロギの存在下では捕食がそれぞれ誘発されました。
クルミを入手する経路に電気ショックの焼き網を配置すると、クルミの摂取量は減少しました。
しかし水道周囲灰白質の腹側部に刺激を受けたマウスは、焼き網が無い状態に匹敵するクルミの摂食量をみせました。
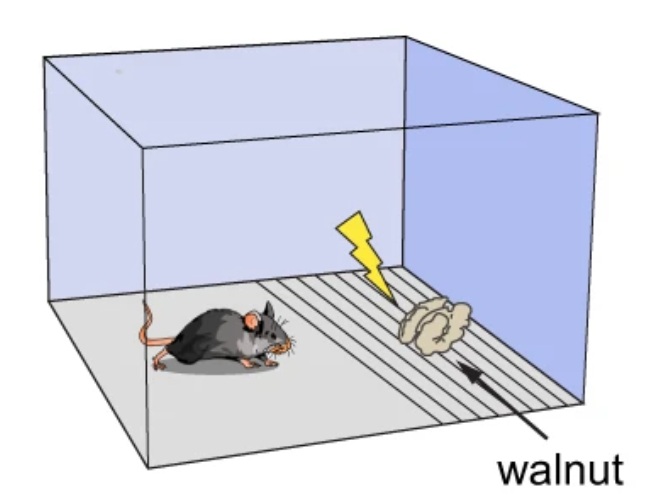
また、長い棒にボールを取り付け、「BG」の文字を綴る軌道でボールを動かすと、水道周囲灰白質に電気刺激を受けたマウスは執拗にボールを追いかけました。
刺激を受けなかったマウスではこのような行動はみられませんでした。
このボールへの追尾は、水道周囲灰白質のレーザー刺激によって誘発された、コオロギの執拗な追跡によく似ていました。
飲水
外側視床下部
アメリカ国立がん研究所では、ラットの外側視床下部への電気刺激で飲水が生じることが発見されました。(Greer 1955)
刺激を受けたラットが、激しい「舐め」行動への強い衝動に駆られていることが、すぐにわかりました。
ラットは後肢で立ち、ガラスで囲まれた円形ケージの周りを勢いよく走り回り、ガラスの壁を激しく舐めました。

この行動は、刺激を止めるとすぐに止まりました。刺激強度を徐々に上げると、舐める行動は徐々に激しくなりました。
水ボトルがある状態では、舐める行動は飲水行動に変化しました。刺激がオンになるとすぐに、ラットは水ボトルに向かって飛びつき、スイッチがオフになるまで貪欲に飲み続けました。
水ボトルの水道水を生理食塩水に置き換えると、ラットは以前と同じように激しく飲み始めますが、生理食塩水がかなり不快であることは明らかでした。
ラットは前肢で注ぎ口を押し、ようやく数秒間ボトルから離れることができましたが、その後再び飲もうとする衝動が強くなり、生理食塩水を舐めることを強いられました。
30分ごとに5分間の継続的な刺激を受けたラットは、毎日0.4 Lを超える水道水を飲みました。
イェール大学のニール・ミラー博士は、外側視床下部の電気刺激によって、苦味剤を混ぜたミルクの飲用をラットに強制できることを示しました。(Tenen and Miller 1964)
まずラットが満足するまで餌と水、そしてミルクを与えた後、外側視床下部への電気刺激により、ラットが再びミルクを飲み始めることを確認しました。
一般的に、すべてのラットは15秒間の刺激の間ずっとミルクを飲み続け、刺激の終了とともに飲用を止めました。
続いてミルクに苦味剤のキニーネを混ぜました。
すると外側視床下部の電気刺激により、ラットに苦いミルクの飲用を強制させることができました。
キニーネ濃度が高いと、一口舐めた後に飲用をやめることもありましたが、数秒後に再び戻ったり、チューブのそばに立って頭と口を使って部分的な「接近-離脱」運動をしたりしました。
キニーネの濃度が上昇するにつれてラットは飲用時間の短縮がみられましたが、刺激の強度を上げることでこれを克服することができました。
最大刺激強度では、ラットは84時間の絶食下で許容したキニーネ濃度の8倍の濃度を許容しました。
カナダの研究者も、外側視床下部の刺激で激しい飲水が誘発されたことを示しました。(Mogenson and Stevenson 1966)
まずラットの外側視床下部に電極を設置し、レバーを押して自己刺激できるようにしました。
12匹のうち10匹のラットは積極的に自己刺激を行い、30分間で4500~5500回レバーを押しました。
続いて自己刺激のレバーを外して給水機を設置して、同じ視床下部外側の電極への実験者による刺激が行われました。
ラットは餌と水を十分に与えられた状態でした。
すると4匹のラットが刺激を受けると水を大量に飲みました。舐め回数は8分間で745~1982回にもおよび、総飲水量は7.5~12.5 mLでした。
残りのラットは刺激に影響を受けず、飲水量は0 mLでした。

飲水行動は刺激に縛られており、電気刺激が終了すると停止しました。
その後1時間で、飲水ラットは4.0~8.5 mLもの尿を排泄しましたが、非飲水ラットは0~0.5 mLの尿しか排泄せず、脳刺激中の水分摂取量が自発的な必要量を超えていたことが確認されました。
続いて自己刺激レバーと給水器を同時に利用できるようにしました。
すると上述の4匹の飲水ラットで、レバーを押しと飲水が交互に繰り返されました。飲水行動は刺激に縛られていましたが、ラット自身が制御するようになりました。
レバーを押すたびに、ラットは電気刺激が終了するまで飲水し、次にレバーを押して自己刺激装置を再びオンにしました。
腹側被蓋野
腹側被蓋野への電気刺激でラットに飲水が生じることも確認されています。(Waldbillig 1975)
反応は視床下部から誘発されたものと類似していました。
1回の飲水は数回の舐めに限られており、これは刺激によって同時に引き起こされた、後退およびしゃがみ動作によって妨害されたためのようでした。
視床下部で誘発される反応とは異なり、腹側被蓋野から誘発される飲水は、刺激の終了後も持続することが多かったです。
扁桃体
扁桃体の電気刺激では、ラットの飲水を誘発するだけでなく、飲水を促進もすることが示されています。(Singer and Montgomery 1969)
この実験では電気刺激の代わりに、神経伝達物質であるアセチルコリンの類似体を扁桃体の皮質核と外側視床下部に注入するという化学刺激が行われました。
まず水を十分に補給したラットにおいて、外側視床下部の刺激によって摂水量が増加することが確認されました。
それに加えて扁桃体の皮質核への刺激も加えると、摂水量の増加が強化されました。
扁桃体への刺激だけでも摂水量の増加が確認されましたが、外側視床下部に比べると程度は小さいものでした。
飲水量
扁桃体の刺激は視床下部外側の刺激で誘発される飲水を促進した。
その他の原始的行動
外側視床下部
ラットの外側視床下部の電気刺激で、かじり行動や巣作り行動が生じることが示されています。(Roberts and Carey 1965)
まず刺激がない状態では、ラットは自発的に板をかじらないことが観察されました。
外側視床下部のいくつかの部位に刺激が加えられると、ラットは探索運動を行い、嗅ぎ回ったり周囲を見回したりしながら、かじり板の縁に遭遇しました。
ラットは切歯で縁を掴み、前肢で隣接する面をつかみ、頭と肩で後方に引っ張りました。破片や欠片が外れるとすぐに口から吐き出され、ラットは再び縁を噛みはじめました。
刺激が止められると、かじりはすぐに止まりました。
空腹なラットですら、刺激がオンになるとすぐに食べるのを止め、板をかじり始めました。板が存在する間中、ラットは食事に戻りませんでした。
板を取り除くと、ラットは探索運動に切り替え、食事は再開しませんでした。
別の部位を刺激すると、口の中に紙片を集める、繰り返し噛んで紙片をずたずたに裂く、鼻先で紙片の山を押す、穴を掘るといった、巣作り行動の要素がラットに誘発されました。
かじる動作と巣作り行動が、同じ部位の刺激によって誘発されることもありました。
ラットの外側視床下部への電気刺激で貯蔵行動が生じることも示されています。(Herberg and Blundell 1967)
実験用ラットは継続的に供給される食物を蓄えることはありませんが、数日間断続的に食物を断つスケジュールに置かれると、満腹時に空腹時と同じ量の食物を蓄え始めます。
実験では、巣材と給水ボトルを備えた、部分的に囲まれた家をケージごとに設けました。
家の外に100個の餌ペレットと同数の類似の木片を置いて、10分間に家の中に持ち込まれた餌ペレットの数を数えました。
外側視床下部の電気刺激が無い場合、満腹ラットは6個程度しか家に持ち込みませんでした。
対照的に刺激が有る場合、満腹ラットは即座に持続的な貯蔵行動を始め、多数のペレットを家に持ち込みました。
貯蔵されたペレットの数は26個程度と、餌が剥奪された時のものに匹敵しました。
ラットは各刺激試験のほぼ半分をペレットの運搬に費やし、残りの時間では集めた量よりはるかに少ない、2個程度のペレットしか食べることができませんでした。
ラットの外側視床下部の電気刺激で物運びが生じることもを示されています。(Phillips et al. 1969)
げっ歯類は、貯蔵、巣作り、および仔獣を運ぶ状況で物体を運びますが、石など明らかな用途のない物体を運ぶこともあります。モリネズミは、より魅力的な光沢のある物を求めて、運んでいた物を置き去りにすることさえあります。
実験では光センサーを使って、ラットがケージの半面に入ると外側視床下部の電気刺激がオンになり、もう半面に入るとオフになるようにしました。
ラットをケージに入れると、数分以内に部屋を行き来して自己刺激することを学習しました。
その後、ケージのオン側に、餌のペレット、ダボピン、ペレットと同じ大きさの木片、消しゴムなどの物体を山積みにしました。
するとラットは口で物体を拾い上げ、ケージのオフ側に運び、刺激がオフになるとすぐにそこに置きました。
ラットが刺激を受けていないときは、物体は決して拾われませんでした。
ラットは非食用物よりも餌ペレットを多く選択しましたが、かなりの数の非食用物も持ち運ばれました。10分間の平均は、餌ペレット40個と非食用物26個でした。
腹側被蓋野
腹側被蓋野への電気刺激でもかじり行動が生じることが示されています。(Waldbillig 1975)
腹側被蓋野から得られる齧り行動は、外側視床下部から得られるものに似ていました。
これは木の縁を切歯で掴み、前肢で隣接する面をつかみ、頭と肩で上方および後方に引っ張るという動作でした。
破片や断片が抜けるととすぐに口から吐き出され、ラットは再び縁を噛みました。
突き出た金属の端に向かって噛むことも時々ありましたが、持続することはありませんでした。
視床下部で誘発される反応とは異なり、腹側被蓋野から誘発されるかじりは、刺激の終了後も持続することが多かったです。
衝動性・多動性
側坐核
オランダの研究者は、側坐核の電気刺激でラットの衝動性が増加することを示しました。(Sesia et al. 2008)
この実験では、ラットはケージ内に設置されたパネルを、音が提示されるまで押し続けなければなりませんでした。
音が提示された後にレバーが挿入され、ラットはレバーを押すことで餌のペレットを得ることができました。
パネルを押してから音の開始までの時間は、各試行において0.6秒から1.5秒の間で無作為に選択されました。
側坐核シェルに電気刺激を受けたラットは、音が提示されるまでパネルを押し続けることができず、餌の獲得に失敗することが多くなりました。
これはラットの衝動性の増加とみなされました。
一方で、側坐核コアに電気刺激を受けたラットは逆の傾向を示しました。
腹側被蓋野
腹側被蓋野と側坐核の電気刺激でネコが多動になることが示されています。(Goldstein and Siegel 1980)
腹側被蓋野へ電気刺激を与えると、ネコは頭で探索するような動きを示しました。
刺激強度を増加させるとネコは多動になり、持続的に歩き、ミャーと頻繁に鳴きました。
側坐核への電気刺激では、ネコはわずかに身をかがめました。
刺激強度を増加させると、ネコは大きく身をががめ、排尿し、そして顕著に多動になりました。
1匹のネコは刺激の終了後に、極度の多動、大きな鳴き声、排尿、および大量の唾液分泌という、発作前のような行動を示しました。
