激怒

脳の特定部位の電気刺激により、人間や動物に激怒、攻撃行動を引き起こすことができます。
よく知られた部位は扁桃体、視床下部、水道周囲灰白質の3つです。
これを悪用すると、暴力的傾向のない個人を暴走させて暴力犯罪を犯させることも可能になると思われます。
少なくとも実際に、遠隔から無線で被験者に暴力行動を誘発する実験が、50年以上も前にすでに行われていました。
目次All_Pages
回路
激怒
激怒回路は大脳新皮質の下にある大脳辺縁系と中脳に存在し、人間と他の哺乳類で相同であると考えられています。 (Panksepp 1998)
これは扁桃体から視床下部を通り、中脳の水道周囲灰白質まで走っています。
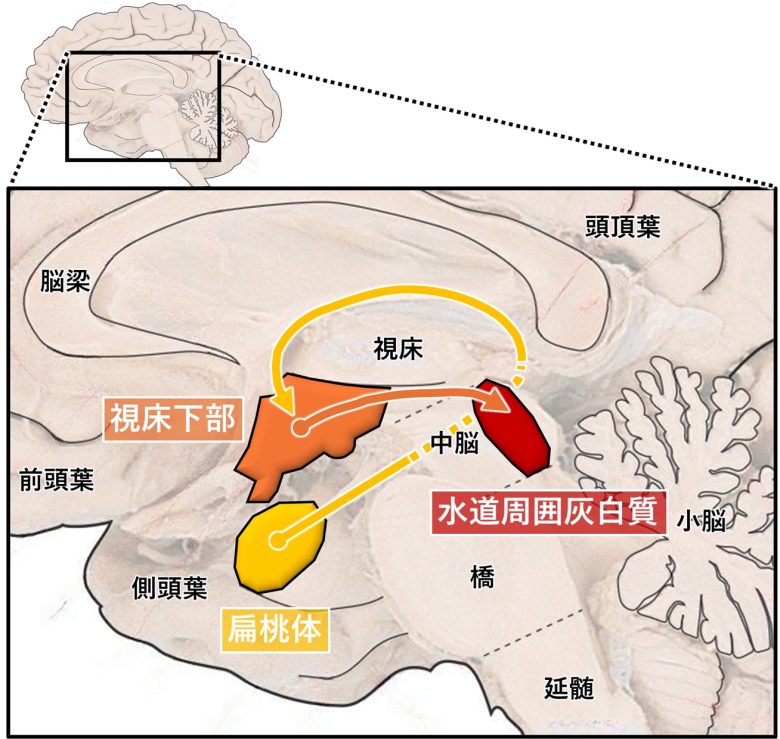

激怒に関する3つの主要な脳部位。扁桃体と視床下部と水道周囲灰白質。
(Panksepp 1998を改変、脳画像はVanderah 2018から)
これら3つの領域の電気刺激で、人間・動物の双方で激怒反応が生じることが確認されています。
大脳皮質を意思決定中枢、視床下部と中脳を実行中枢とするならば、扁桃体はその中間に位置する構造です。(Haller 2018)
一般的に、扁桃体が大脳皮質による抑制から解放され、視床下部と中脳の水道周囲灰白質にある実行領域を活性化することで激怒が生じると考えられています。
これらの領域は、上位にある構造が下位の完全性に依存するように階層的に配置されています。(Panksepp 1998)
つまり、水道周囲灰白質が損傷していると、扁桃体あるいは視床下部の電気刺激で激怒反応は生じなくなります。(de Molina and Hunsperger 1962)
しかし扁桃体あるいは視床下部が損傷していても、水道周囲灰白質の電気刺激で依然として激怒反応が生じます。
攻撃
攻撃は怒りよりも幅広い現象で、攻撃に常に怒りが伴うとは限りません。 (Panksepp 1998)
攻撃のタイプは主に防衛的激怒と捕食攻撃の2つのカテゴリに分類することができ、動物が激怒反応を示すのは前者の反応のみです。
この2つの攻撃に関しては、明確な神経回路の分離がみられます。
防衛的激怒はライバル攻撃とも呼ばれ、扁桃体の内側核、視床下部の腹内側核 (※) 、水道周囲灰白質の背側部が関与しています。(Haller 2013, Haller 2018)
正確には視床下部の内側基底部。
捕食攻撃には扁桃体の中心核、視床下部の外側部、水道中心灰白質の腹側部が関与しています。
またこのほか、腹側被蓋野も捕食攻撃に関与しています。(Siegel and Victoroff 2009)
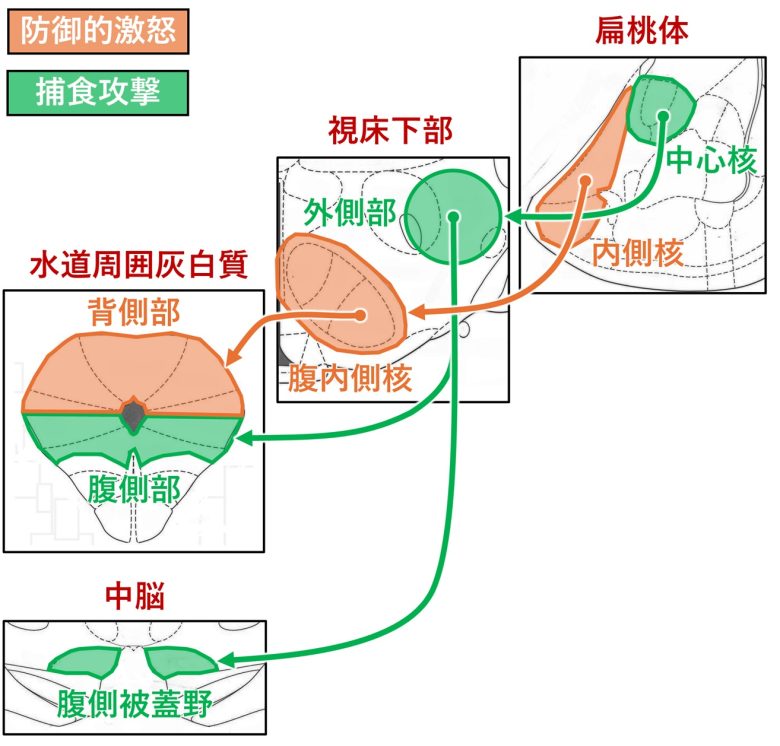
防衛的激怒と捕食攻撃の神経回路
(Haller 2018を改変、脳地図はPaxinos and Watson 2009から)
一般に「怒り」や「激怒」と呼ばれる感情は、その原動力と感情的な影響の多くを、防衛的激怒の神経回路から得ている可能性が高いです。 (Panksepp 1998)
防御的激怒では交感神経が活性化し、反応は非常に衝動的で、理性を司る大脳皮質の関与を欠いています。(Siegel and Victoroff 2009)
この行動は自然界において、同種・異種の他の動物による脅威に反応して起こります。
一方、捕食攻撃では交感神経の活性化はほとんど見られません。またこの反応には計画と戦略が必要であるため、大脳皮質が関与していると考えられています。
動物ではこの行動はほとんどの場合、異種の動物、通常は典型的な獲物 (ネコがネズミを攻撃するなど) に向けられます。
捕食回路は欲求回路の一部と考えられており、実際この回路を刺激すると、ある種では摂食につながり、他の種では捕食的な追跡につながります。(Panksepp 1998)
欲求回路に関しては、詳しくは4ページの「快楽と欲求」をご覧ください。
人間の捕食攻撃
人間の捕食攻撃のいくつかの特徴は、動物にみられるものと類似しています。(Meloy 1988)
交感神経の覚醒が最小限であり、対象は脅威として認識されず、行動が計画的で目的を持っており、攻撃対象が他に移らないことなどです。
しかし、動物では捕食攻撃は通常は異種に向けられるのに対し、人間では同種に対して向けらることがあり、様々な目的によって刺激される可能性があります。
- 復讐心の充足
- 強迫的な衝動からの解放
- サディスティックな欲望の充足
- 偏執性妄想や幻聴などの精神病症状の緩和
- 病的な嫉妬や羨望の感情の緩和
連続殺人や大量殺人においても、捕食攻撃の神経回路が使用されている可能性があります。
人間
扁桃体の内側領域の電気刺激で、人間が激怒し、攻撃行動を起こすことが示されています。
これは動物で「防衛的激怒」を誘発した領域に該当します。
また中脳の水道周囲灰白質の腹側部の刺激で、殺意さえ表明する強烈な激怒と攻撃行動が生じた例があります。
これは動物で「捕食攻撃」を誘発した領域に該当します。
ヒースの人体実験
精神科医のロバート・ヒースは1949年にテュレーン大学に精神・神経科を創設し、そこで統合失調症の患者を対象にした脳電気刺激の人体実験を行っていました。(Heath 1996)
ヒースは中脳被蓋、視床下部、内側扁桃体、海馬の各部位を電気刺激すると、さまざまな強さの恐怖や怒り、時には暴力の反応が引き起こされた、と述べています。
水道周囲灰白質
男性患者「A-10」には、中脳被蓋、おそらくは水道周囲灰白質の腹側部 (※) への電気刺激が加えられました。(Heath 1954, Heath et al. 1954)
p44の「中脳被蓋の電極は水道から数ミリ腹側 ( a few millimeters ventral) 」という記述と、p560の「今のところ患者A-10のみが中脳被蓋の刺激を受けた」という記述と、下記の刺激が動眼神経を活性化したという記述から。
すると患者に激怒が誘発され、居合わせた医者に突進し、お前を殺すと殺意さえ表明しました。
この様子をカメラで撮影した記録がテュレーン大学には残っていました。(Berns 2005)
電気刺激が始まると、患者の眼球が上下に揺れ動きました。(※)
これは刺激が動眼神経を活性化したためでした。(Heath 1992)
彼は蜂に刺されたイヌのように頭を数回振り、てんかん発作を起こしたように見えました。しかし意識は完全にありました。
彼は複視と身体全身の強い不快感を訴え、右手でかぎ爪を作り、胎児のような姿勢で丸まりました。
刺激が続けられ3分間を過ぎると、彼はますます狂乱的になり、「脳がこんな風になっていると何も考えられない。ああ、だめだ……気絶する前に!気絶したくない……ああ、俺の脳が!」と叫びました。
突然、患者の声が変わり、理解できないほど高い声で叫びました。
それから彼は服を引き裂き始め、シャツを剥ぎ取ろうとし、担架から立ち上がりました。
面談者が「服を引き裂いていますね。自分でも服を引き裂いていると気づいていますか?」と聞かれると、患者は支離滅裂になりそうになりながら裏声で、「構わねえ!何かしねえと!構わねえ!構わねえ!」と叫びました。
少しの間止まり、彼は再び担架から降りようとし、「お前を引き裂いてやる!」と叫びました。
数人の手が現れ、患者を押さえつけ、手を縛りました。
「やめなさい!」と面談者が命じました。「やめなさい!」
患者はカメラを見つめ、叫びました。「知ったことか。お前を殺してやる。起こせ。お前を殺して、ずたずたに引き裂いてやる!」
刺激が止められると、患者は自分の反応に困惑しました。「彼に恨みはありません。ただ、一番近くにいただけです」と語りました。(Heath 1996)
内側扁桃体
27歳の女性患者「A-8」には、扁桃体への電気刺激が加えられました。(HEATH et al. 1955, Heath 1996)
電極は外側扁桃体に配置され、これは内側扁桃体の近隣にありました。
そこへの低強度の刺激は快楽反応を誘発しました。
対照的に、刺激電流を内側扁桃体まで広がり活性化するほど増大させると、激怒や暴言を特徴とする嫌悪反応が観察されました。
しかし、刺激電流を減少させ、内側扁桃体が活性化しなくなると、患者は明らかな喜びとともに反応し、しばしば笑い、そして声に出して「なぜそんなことをしたのだろう」と不思議がりました。
以下は電気刺激を加えられた時の面談の様子です。(King 1961)
刺激前の患者の声の調子は非常に平坦で、顔も無表情で変化がありませんでした。
彼女は「すべてが間違っている。私は何の一部でもないような感じ。私がどこにも属していないように、すべてが夢かなんかみたい」と離人感や現実感の消失を訴えていました。
ここで5 mAの電気刺激を扁桃体に加えられました。面談者がどんな気分かと尋ねると、彼女の声の調子が非常に高くなりました。
彼女は「この椅子から立ち上がりたい!そうはさせないで!」と叫び、表情が一変して嘆願する様子を見せました。
「こんなことしないで!私は意地悪になりたくない!」
面談者が自分のことを殴りたいかと聞くと、彼女は「ええ、何か殴りたい気持ちよ。何かつかんで、引き裂きたい。そうしないように、取り上げて!」と答え、彼女は自分のスカーフを面談者に手渡しました。
彼は代わりに彼女に紙の束を手渡すと、彼女はそれ以上言葉を交わすことなく、それをずたずたに引き裂いてしまいました。
「こんな風に感じるのは嫌よ!」
ここで電流が4 mAまで下げられました。すると彼女は即座に大きな笑顔を浮かべ、少し気分がよくなったと言いました。
彼女に先ほどの事について聞くと「この椅子から立ち上って、出ていきたかった。なにかを殴りたかった。何かを破りたかった。何でも。あなたじゃない、何かよ。立ち上がって、破きたかった。自分を制御できなかったわ。」と答えました。
ここで再び、電流の刺激レベルが5 mAまで上げられました。彼女の声は大きくなり、懇願するような調子になりました。
「私にあなたを殴らせないで!」
再び気分について聞かれると、「少しだけ気分がいいかもしれないわ。すべてを吐き出すから。こうなってるとき、他の考えが浮かんでこない。血圧をとって!これを切って!こんなの辛すぎる!血圧をとって!」と彼女は怒りの表情を浮かべて強い抑揚で叫びました。
「ここに閉じ込めるのはやめて!立つわ!私を置いておきたいなら、他のだれか連れてきた方がいいわ!私、あんたを殴るわよ!」
彼女は殴るように腕をあげました。
ここで電流が4 mAまで下げられました。患者は再び大きな笑顔になり、笑い声をあげました。
「なぜ私をこんな風にできたの?どうすることもできなかった。あなたの顔をひっぱたいてやりたかった。あんな風にされるのは嫌い。」
アービンとマークの人体実験
内側扁桃体
ハーバード大学の精神科医フランク・アービンと神経外科医バーノン・マークは、マサチューセッツ総合病院で暴力的傾向のある多数の患者を研究していました。(Scheflin and Opton 1978, Breggin 1994)
彼らはイェール大学のホセ・デルガドから電波を使った遠隔脳刺激装置であるスティモシーバを供与され、遠隔から無線で被験者に暴力行動を誘発する人体実験を行っていました。
彼らの有名な被験者の一人は34歳の男性患者トーマス・R 氏で、彼は優秀なエンジニアで重要な特許をいくつか取得していました。 (Mark and Ervin 1970)
筋骨隆々な体格にもかかわらず、彼は礼儀正しく同情的で、その態度は物静かで控えめでした。激怒していないときに彼が暴力行為を起こすことができるとは信じがたいものでした。
彼の主な問題は、暴力的な怒りでした。これは同僚や友人に向けられることもありましたが、ほとんどの場合、妻や子どもに向けられました。
妻に対する暴行は、腹部・顔面の激しい痛みを訴えた後に始まるのが典型的でした。
被験者の扁桃体の内側領域に電気刺激を加えると、この痛みの訴えと、「制御を失っている」という感覚が生じました。
これらの反応はいずれも、患者の暴力行為の始まりを示す兆候であることが分かっていたため、怪我する可能性を避けるため、刺激は短時間に抑えられました。
後日、同じ部位を短時間刺激したところ、この前兆的な現象が再現されました。
その領域から4ミリ横にある扁桃体の外側領域への電気刺激は、反対の反応を引き起こしました。
刺激が始まってから 60~90秒以内に、彼の気分は考え込んでいる状態から、くつろいだ状態へと変化しました。
このくつろいだ状態は、電流を切っても消えることはなく、4~18時間かけてゆっくりと消えていきました。
アービンとマークの手掛けたもう一人の有名な被験者は、22歳の女性患者のジュリア・P 氏です。(Mark and Ervin 1970, DELGADO et al. 1968)
ジュリアさんは10歳の頃から側頭葉てんかんを患い、頻繁に怒りの爆発を起こし、その回数は計12回に及び、そのうちのいくつかは暴力事件に発展しました。
ある時は映画館のラウンジで、ジュリアさんの左腕と手にぶつかった少女の心臓を突き刺しました。また別の時には、病院で発作の兆候を見せたジュリアさんを助けに駆けつけた看護師を、ハサミで肺に刺しました。
彼女の右の側頭葉にスティモシーバが装着され、内側扁桃体に遠隔からの無線刺激が加えられました。
すると彼女は唇を引っ込めて歯をむき出しにする、一連の怒りの表情を作りました。太古から伝わる「霊長類の威嚇ディスプレイ」でした。
続いて壁に向かって飛びかかり、攻撃しました。
これは突然で予期せぬ出来事で、なぜ彼女の攻撃の被害者は身を守る時間がなかったのかがわかりました。
2回目の刺激は、ジュリアさんがギターを弾いている時に始まりました。刺激が始まって5秒後、ジュリアさんは歌うのをやめ、ぼんやりと前を見つめました。会話から遠ざかり、彼女は診察していた精神科医の質問に答えることができませんでした。
すると突然、彼女はギターを突然力強く投げつけました。ギターは精神科医の頭をかすめ、壁に激突しました。
彼女は数分間部屋の中を歩き回った後、徐々に静かになり、彼女のいつもの陽気な振る舞いを再開しました。
トーマス・R 氏とジュリア・P 氏、両名ともに後に扁桃体の内側領域の破壊術を受け、暴力的な行動が鎮静されたと、アービンとマークは報告しています。
しかしこれは家族や病棟の看護師から手紙を受け取って、この件を独自に調査を開始したハーバード大学卒の精神科医ピーター・ブレギン博士の主張とは異なります。
ブレギン博士は、扁桃体の破壊後もジュリアさんは衝動性が無くならず、衰弱し始めて自殺願望を抱き、いまだ入院したままであることをを突き止めました。
トーマスさんに関しては、そもそも彼は暴力的な人物ではなく、脳電気刺激の実験に不本意に参加させられ、本来不必要であった脳外科手術によって人格を破壊されてしまったことがわかりました。
彼の暴力行動は、アービンとマークの介入が始まってから起きたものでした。
詳しくは10ページの「黎明期のマインドコントロールの試み」をご覧ください。
特に暴力的な問題のない、末期の扁平上皮癌を有する、63歳の温厚な機械工も内側扁桃体への電気刺激を受けました。(Stevens 1969)
電気刺激は疼痛軽減を目的とした、扁桃体破壊術の術前に行われました。
内側扁桃体の電気刺激後、彼は目を荒くし、看護師が近づくのを許さず、呼吸が荒くなり、非常に怒っているように見えました。
虚ろな視線を送り、ベッドで立ち上がり、近づく者を攻撃し、便を失禁し、その便は彼に近づく者にかなりの成功を収めて投げつけられました。
この行動は、看護師と病棟スタッフの一団が彼を拘束するまで約10分間続きました。
翌日、電気刺激が前回より短い時間行われ、再び同様の、しかし前日より軽い発作が起こりました。
動物を使った激怒・攻撃行動の誘発実験では、同時に失禁が起きる事はしばしば観察されています。
扁桃体の破壊によって痛みは軽減されず、電極埋込みから7週間後に患者は死亡しました。
その他の研究
内側扁桃体
イギリスのエディンバラ大学では、暴力行動の治療として、内側扁桃体の凝固術が適用されました。(Hitchcock and Cairns 1973)
その術前に、18人の患者のうち6人は内側扁桃体の電気刺激も受けました。
刺激の最も顕著かつ劇的な効果は、さまざまな攻撃的な反応を引き起こしたことでした。
反応は、適切に差し向けられた筋が通った言動から (外科医に向かって「立ちあがってお前に噛みつきたい」と話す)、制御不能な罵声や物理的な破壊行動まで、様々でした。
4人の患者においては、カーテンや衣服を引き裂いたり、定位固定フレームに手を伸ばして無理やり外そうとしたりするような、落ち着きのない、似通った行動パターンがみられました。
彼らの行動は口頭の指示によって止めることはできませんでした。
2名の患者では、定位固定フレームが患者によって外されるのを防ぐために、力ずくで対処する必要がありました。
これらの発作の前後では、この2名の患者は状況を十分に理解しているように見え、フレームに触れたり、手術を妨げたり、カーテンなどをいじったりすることを一切控えていました。
その他、3人の患者が逃げ出したいという欲求を示しました。
刺激開始から3秒以内に反応が見られたものもあれば、刺激停止後1分経っても持続するものもありました。
激しい罵倒が生じた2件のうち1件では、罵倒は刺激から30~45秒後に突然始まりました。これは3回の機会で確認されました。騒乱は15~30秒間続き、刺激停止から5~30秒後に患者は落ち着きを取り戻しました。
ある時、刺激を止めてから30秒後に、患者に怒りを感じたかどうか尋ねました。彼は怒っていたことは認めましたが、今はもう怒っていないと答え、非常に驚いた様子でした。
ある患者では、埋め込んだ電極を除去する際に受けた損傷の結果、重度の右半身麻痺が生じ、片腕に重大な障害が残りました。
視床下部
パリのサルペトリエール病院では、57歳のパーキンソン病の男性に、治療として視床下核への電気刺激が施されました。(Bejjani et al. 2002)
有効な電極接点の探索中に視床下部の後内側部の接点が刺激されると、数秒後に突然の攻撃行動の発作が生じました。
患者は興奮して激怒し、叫び始めました。患者は定位固定フレームを外そうとしながら手術台から降りようと激しく試みたため、強制的に拘束しなければなりませんでした。
約5分間続いたこの発作の間、患者は緊張した表情を浮かべ、眼球が突出し、頻脈を示しました。
彼は警戒心が強く、混乱や発作の兆候は見られませんでしたが、攻撃的な発作の激しさのため、これを裏付けることはできませんでした。
刺激は直ちに中止され、鎮静剤が注射されました。患者が鎮静から覚醒した後、攻撃的な行動は再発しませんでした。
患者は、自分が攻撃的だったことと「何かがおかしかった」ことを思い出しましたが、行動を説明することはできませんでした。
有効な視床下核の電極接点への電気刺激は運動障害を81%改善し、投薬量を68%減らすことができました。
ネコ
ネコは激怒・攻撃行動の研究でよく利用されてきた動物です。
これらの研究から、激怒・攻撃行動に扁桃体、視床下部、水道周囲灰白質の3つの領域が主に関与していることが明らかになりました。
視床下部
スイスのチューリッヒ大学の生理学者、ヴァルター・ヘス博士は1930年代に間脳 (視床と視床下部) の機能をマッピングする研究を始め、1949年にノーベル生理学・医学賞の受賞者しました。
博士はその一連の研究の中で、ネコの視床下部の電気刺激により、激怒および攻撃行動が生じることを発見しました。(Hess and Brügger 1943, Hess 1958)
刺激を受けたネコはまるでイヌに威嚇されているかのような行動をとりました。
ネコは「カッ!」と怒声を浴びせ、鼻を鳴らして唸り声を上げ、背中の毛を逆立て、尻尾をふさふさにし、瞳孔は開き、時には最大限にまで開き、耳は後ろに倒れたり、前後に動いたりしました。
刺激が持続または強まると、ネコは実際に攻撃を仕掛けました。近くに立っている人の方を向き、飛びかかったり、前足で狙いを定めて一撃を加えたりしました。
このような興奮した状態で、ネコが糞を排出することも珍しくなく、より頻繁には尿を噴出させました。
これらの行動は電気刺激が停止したとたんに終わりました。しかし、数分間はネコはイライラした様子で、研究者が近づくとネコは敵意を示しました。
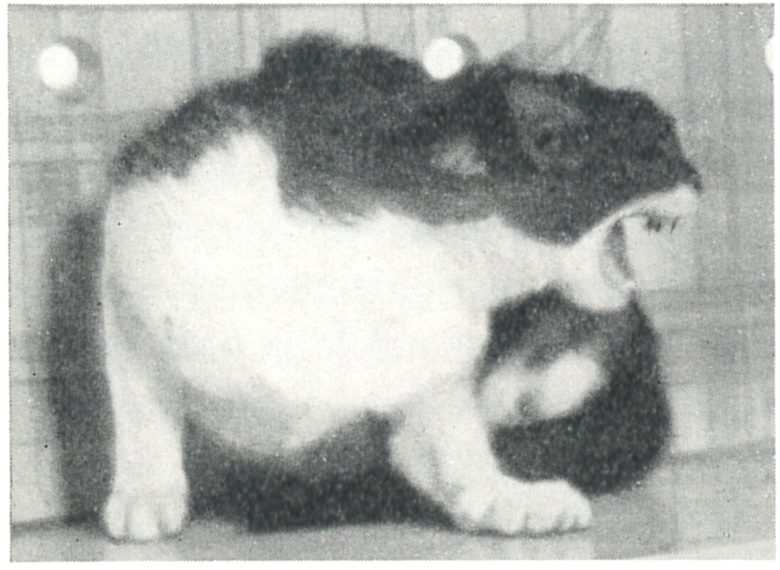

別の研究から。視床下部の電気刺激で、感情的な防御反応が誘発された。(Akert 1961)
それから数多くの研究者がヘス博士の実験に触発され、脳の電気刺激による激怒と攻撃反応の誘発実験を行いました。
イェール大学の神経生理学者ホセ・デルガドもその一人です。(Delgado 1964)
ネコの視床下部を電気刺激すると、瞳孔が広がり、毛が逆立ち、シャーと鳴き、歯をむき、唸り声をあげるなどの攻撃的な態度を見せました。
ネコはその後前進し、爪を立て、狙いを定めて他のネコに打撃を与え、攻撃を受けたネコは反撃するか、退きました。
研究者が攻撃されたネコを守ろうとすると、刺激を受けたネコの攻撃はそらされ、手袋をした手を凶暴に噛み、引っ掻きました。
攻撃行動は通常は刺激とともに始まり、刺激とともに終わりましたが、実験ネコに対する不信感が残りました
たとえ刺激を受けたネコが小さく、他のネコに殴られたり引っかかれたりしても、視床下部が刺激される度に攻撃や戦いが引き起こされました。痛みを伴う経験は引き起こされた攻撃性を抑制しませんでした。
またネコはこの刺激を嫌がっており、ペダルを回すことにより脳刺激を止めることを学習しました。




視床下部の腹内側核
後続の研究者は、視床下部で激怒・攻撃反応が生じる部位をより詳細に調査しました。
ワシントン大学の研究では、ネコの視床下部の中間部、特に腹内側部の電気刺激で攻撃行動が生じることが示されました。(Nakao 1958)
典型的な反応として、瞳孔の散大、耳の後退、背中の反り、脚の伸展などがありました。
弱い電気刺激では、ミルクを飲む、毛づくろいをする、喉を鳴らす、歩くなどの自発的な行動が突然停止しました。
そして瞳孔が開き、わずかに唸りましたが、動くものや実験者が視界に入らなければ、反応はそれ以上進みませんでした。
しかし、実験者が見えると、ネコは耳を引っ込め、背中を丸め、実験者に向かって歯を剥いてシャーと鳴きました。
より強い電気刺激では、実験者が少しでも動くと、方向づけられた激しい攻撃を引き起こしました。棒を差し出すと、ネコは激しく噛みつきました。
日本の九州大学の研究では、ネコの攻撃行動のほとんどが視床下部の中間部にある、腹内側核への電気刺激で生じました。(Yasukochi 1960)
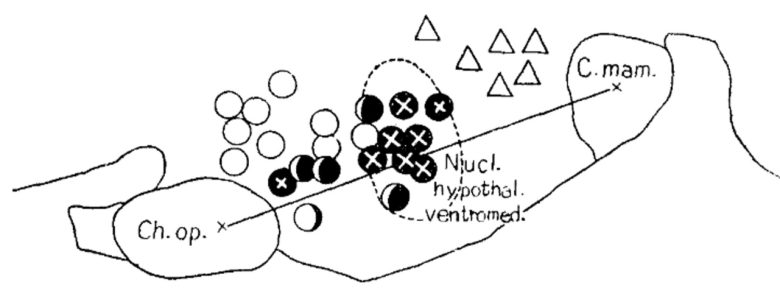
黒丸:不快な苛立ち・激怒、黒丸にバツ:激怒および攻撃、白丸:不安・恐怖、△:渇望・切望。
ネコは刺激が弱い間は、ただ辺りを睨みつけ、不快そうに唸り声を上げていました。
刺激の強さが増すにつれてネコは徐々に興奮して攻撃的になり、瞳孔は大きく開き、耳は後ろに引っ込み、毛は逆立ち、威嚇するように歯をむいてシャーと鳴き、ついには挑発する実験者に飛びかかり、怒りの唸り声を上げながら襲い掛かりました。
一部のケースでは、まるで何らかの敵対的な物体を幻視したかのように、ネコがケージの無害な角に激しく攻撃を仕掛けることもありました。
水道周囲灰白質
激怒・攻撃反応は中脳の水道周囲灰白質の電気刺激でも生じることが示されています。
チューリッヒ大学のロベルト・フンスペルガ博士は、ネコの水道周囲灰白質への電気刺激で激怒と攻撃反応が生じることを発見しました。(Hunsperger 1955)
刺激に対する怒りの反応は、以下のように進行しました。まずまぶたが開き、瞬膜が後退し、瞳孔が散大しました。
その後、興奮状態に入りました。瞳孔はさらに散大し、時には最大限にまで開きました。ネコは目に見えない敵をじっと睨みました。突然、歯をむいてシャーと繰り返し鳴きました。この爆発的な行動には、唸り声も伴いました。
刺激が強まると、ネコは目の前の人や棒切れを攻撃するようになりました。
これらの激怒・攻撃反応は、視床下部よりも水道周囲灰白質からの方がより早く出現しました。
ネブラスカ大学の研究者もネコの水道周囲灰白質の電気刺激で激怒と攻撃反応が生じることを示し、フンスペルガ博士の発見を裏付けました。(SKULTETY 1963)
刺激を受けたネコは、一般的に怒り反応として、耳を平らにし、背中を反らせ、背中と尾の毛を逆立て、瞳孔を広げ、唸り声を上げシャーと鳴き、頻繁に排尿するなどの特徴を示しました。
観察箱に手袋を入れると、ネコは狙いを定めて攻撃を仕掛けました。
刺激を止めるとすぐに反応は止まりましたが、ほとんどのネコはその後、唇を舐め、ニャーニャーと鳴き、15~30秒間落ち着きなく歩き回りました。
扁桃体
扁桃体の電気刺激でも激怒反応が生じることが示されていますが、他の部位と比べると攻撃まで至ることは少ないようです。
フランスの神経科医は、ネコの扁桃体の電気刺激でネコに激怒反応が生じることを発見しました。(Gastaut et al. 1952)
弱い刺激では「注意」の態度が生じ、ネコは目を見開き、首と耳をまっすぐに伸ばしました。
刺激が強くなると「恐怖」の態度が生じ、ネコは頭を下げ、匂いを嗅ぎ、耳を後ろに反らせ、隠れようとするように見えました。
さらに強い刺激では「激怒症候群」が生じ、ネコは体を反らせ、耳を引っ込め、毛を逆立て、唸り声を上げました。
怒りは方向性がなく、電気刺激を受けている間、ネコは外部刺激に対して通常より感受性が鈍くなりました。
内側扁桃体
デューク大学の研究では、ネコの扁桃体のうち、内側核と中心核への電気刺激で激怒反応が生じることが示されました。(Shealy and Peele 1957)
刺激を受けたネコは唸り声を上げ、後ろ足で立ち上がり、爪を出し、空を掻き、近くにあるものなら何でも噛んだり引っかいたりしようとしました。
怒りは特定の人物や物に向けられたものではありませんでした。
これに加えて、唾液と鼻水の分泌、嘔吐、くしゃみ、排尿、排便といった、自律神経の反応が誘発されることもありました。
フンスペルガ博士は、扁桃体だけでなく、分界条と呼ばれる神経線維にも焦点をあて、ネコの激怒反応を誘発する部位を詳しく調べました。(de Molina and Hunsperger 1959)
分界条は扁桃体の主要な出力経路として機能する神経繊維の束で、扁桃体の内側部から視床下部の腹内側核まで伸びています。
先述の通り、視床下部の腹内側核は激怒・攻撃反応を誘発する領域です。
激怒反応を誘発した電極は、分界条が起始する扁桃体の内側領域に集中していることがわかりました。これは内側核と中心核に隣接する領域、および基底核の背側部に該当しました。
また分界条自体の電気刺激でも激怒反応が誘発されました。
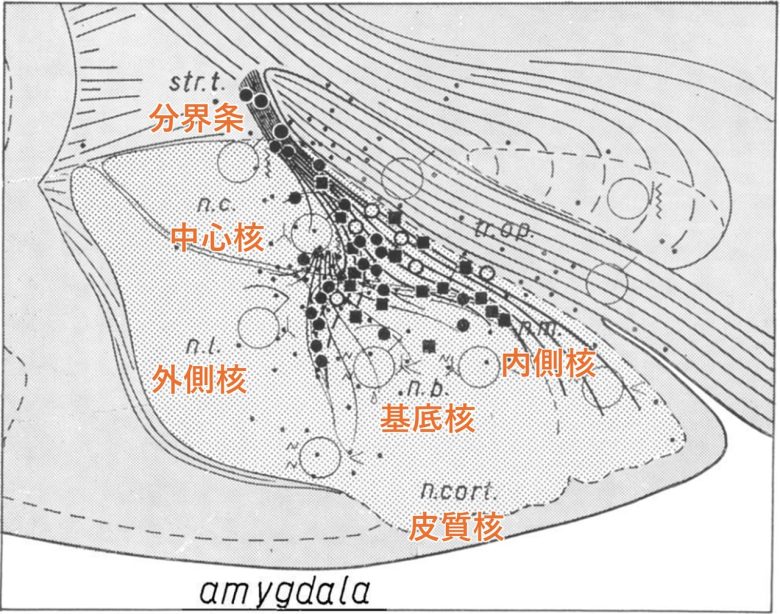
扁桃体と分界条で激怒反応を誘発した部位。黒丸:威嚇・うなり声、黒四角:威嚇・うなり声・シャーという鳴き、白丸 :予期せぬ逃走。(Hunsperger and Bucher 1967)
刺激開始前のネコは、テーブルの上に静かに座るか横たわっていました。
刺激開始直後、ネコは観察者の方に頭を向け、まぶたが開き、瞳孔が散大し、呼吸の深さと速度が増加しました。目は観察者を見つめたままで、ネコは緊張しているように見えました。
次に、低い唸り声を発し、顎を突き出し上唇をすぼめ、続いて5~10秒間隔でより大きな唸り声を発しました。
しばしば瞳孔は最大限に散大し、耳は後ろに倒れ、頭は下げられ、背中と尾の毛が逆立ちました。
唸り声は、時々短い吐息のような呼気と交互に現れ、その間中、ネコは立って背中を少し丸めていました。
誘発された反応が唸り声に続くシャーという鳴きであった場合、刺激の終わりに近づくと、ネコは騒々しいシャーという鳴きを繰り返すようになり、口は大きく開けられ、口角は下がり、歯をむき出しにしました。
ネコは観察者に顔を向けたものの、攻撃することはありませんでした。
刺激部位によっては最終的にネコが逃亡してしまう場合もありました。

部位ごとの違い
フンスペルガ博士は、ネコの視床下部、扁桃体、中脳の水道周囲灰白質の電気刺激がそれぞれ異なる反応を誘発することに気づきました。(Hunsperger and Bucher 1967)
視床下部では、その反応は、ネコがイヌに出会ったとき、あるいは友好的でない仲間のネコに出会ったときに見られる自然な反応と驚くほど似ていました。
ネコは前足を上げ、攻撃態勢を整えました。威嚇の際には、激しいシャーという鳴きも伴いました。時折、激しい威嚇の後、逃げ出すこともありました。
扁桃体では、防御行動に唸り声が伴い、シャーという鳴きは長時間の刺激を受けた後にのみ発生しました。
視床下部から得られる反応とは対照的に、ネコはそこまで感情を顕わにはしませんでした。
中脳の水道周囲灰白質では、常にシャーという鳴きと瞳孔散大が起こりましたが、完全なパターンは確実には生じませんでした。
しかし、刺激してからネコがシャーと鳴くまでの時間は、視床下部よりも短くなりました。



さらに視床下部と水道周囲灰白質の刺激では、棒や人間の手などをネコに近づけると、ネコは攻撃行動に移りました。
視床下部を刺激している間にぬいぐるみのネコをテーブルの上に置くと、威嚇してから時折逃げるという防御パターンが変化しました。
ある時は興奮したネコは、シャーと鳴きながら爪を出し、敵を追い払いました。その刹那、狂乱の中テーブルから飛び降りました。またある時は、シャーと鳴きながら爪を出し、人形の顔面を叩きつけました。



中脳の水道周囲灰白質の刺激で不完全なパターンが生じたのは、統合された攻撃行動を引き起こす地点でなく、その断片的な反応を誘発する地点が刺激されたためである可能性があります。
実際、サルを使った実験で、攻撃行動が中脳含む脳幹レベルで断片的な運動に分解可能であることが示されています。(DELGADO 1967)
防衛的激怒と捕食攻撃
視床下部
イェール大学の心理学者ジョン・フリン博士は、ネコの視床下部の電気刺激により誘発される攻撃行動には、2種類あることを発見しました。(WASMAN and FLYNN 1962)
一つは内側視床下部から誘発される防衛的激怒で、もう一つは外側視床下部から誘発される捕食攻撃です。
防衛的激怒

内側視床下部の電気刺激でラットに向かって激怒するネコ。(Flynn 1967)
この攻撃は、ネコの怒りの兆候であると一般的に考えられている、顕著な交感神経の活性化を特徴としていました。反応は必ず、警戒行動と瞳孔の散大から始まりました。
弱い刺激では、2分間刺激してもネコは警戒しているだけでした。
強い刺激では、最初の警戒に加えて毛の逆立ちが目立つようになり、尾はふさふさ、ふわふわに膨らみました。
シャーという鳴きと低い唸り声が交互に聞こえました。排尿は、セッションの最初の試みでよく見られました。座っていた場合は、ネコは飛び上がりました。頭を地面につけ、背中を反らせ、爪を立て、シャーと鳴きまた唸り声をあげ、大量によだれを垂らし、深く呼吸しながら動き始めました。
ネコはじっと立ってラットをじっと観察しているように見え、感情的な側面はさらに顕著になりました。
1、2秒後、ネコは爪を立て前肢を振り上げ、前肢で素早く正確な一連の連打を加えました。ラットが急に動けば攻撃の引き金になりましたが、ラットが動かなかったり固まっていたりしても攻撃は起こりました。
捕食攻撃
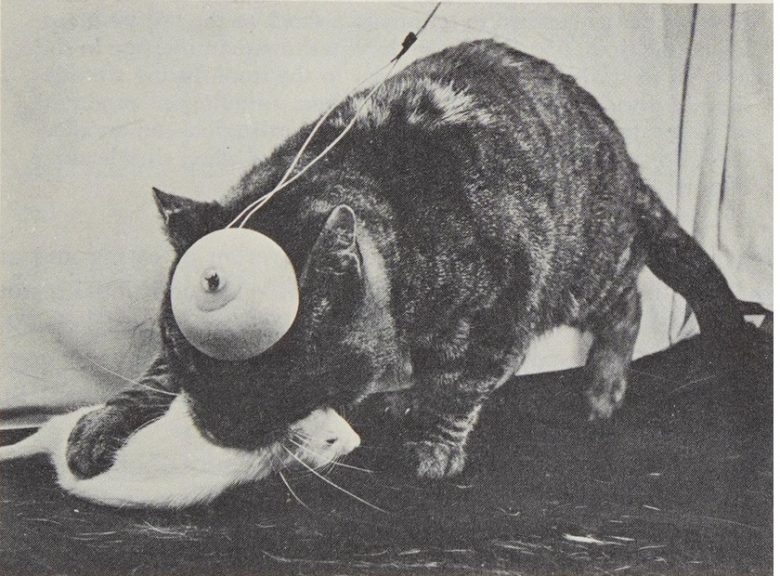
外側視床下部の電気刺激でラットを追跡して首に噛みつくネコ。(Flynn 1967)
この2番目の攻撃は、動物が獲物を追跡する様子を彷彿とさせました。
弱い刺激では、ネコは刺激が始まると通常すぐに飛び上がり、注意深く左右を見回しました。刺激が続くと、ネコはケージ内を歩き回りますが、ラットは無視しました。
強い刺激では、ネコは鼻を地面につけ、背中を反らせ、毛を少し逆立てて素早く動きました。時には、ネコの鼻が軽くピクピクと動き、まるで匂いを嗅いでいるかのようでした。
ネコはたいていラットにまっすぐ近づき、頭と首を激しく噛みつきました。
ネコは前肢を使って主にラットを仰向けに倒し、その喉を狙いました。前肢は散発的な打撃を与えたり、爪でラットを引っかくのには使われませんでした。
ネコが最初にラットの腹や背中に噛みつくこともありましたが、最終的に噛みつくのは頭と首でした。
いくつかの試行では、ネコはラットを無視したり、通りすがりに匂いを嗅いだりしながら、ケージの周りを幾度か周回しました。その最中、ネコは突然ラットに飛びかかり、残忍にもラットを口で引き裂きました。
この捕食攻撃は、特に弱い刺激の場合には、ネコの怒りを示すと思われる兆候が最小限であることに特徴づけられました。
ネコはシャーと鳴くことはほとんどなく、唸り声を上げたり、甲高い叫び声を上げたり、よだれを大量に垂らしたりすることもありませんでした。しかし、ケージ内を動き回るネコの動きはより素早く、より執拗でした。
この捕食攻撃は防御的激怒ほど劇的ではありませんでしたが、実際にはラットにとってはより効果的で、より致命的でした。
防衛的激怒と捕食攻撃の違い
攻撃反応は刺激の消失と同時に停止するという意味で、完全に刺激の制御下にありました。
十分な勢いがあった場合は前足の一撃はそのまま加えられましたが、噛みつきは顎が閉じる直前に停止することができました。
実験には自発的にはラットを攻撃しない柔和なネコが選ばれましたが、脳に刺激を与えると、ラットへの攻撃行動に移りました。
ネコにとって防衛的激怒は嫌悪的で機会があればそれを避けようとしましたが、捕食攻撃に対してはネコは嫌悪反応を示しませんでした。 (Adams and Flynn 1966)
むしろネコにとって捕食攻撃は報酬的であり、通常ラットを攻撃しない柔和なネコが、脳刺激をうけるとラットを攻撃できる部屋に好んで侵入するようになりました。(Roberts and Kiess 1964)
腹側被蓋野
フリン博士は、ネコの捕食攻撃が腹側被蓋野の電気刺激によって生じることも発見しました。(Bandler et al. 1972)
この捕食攻撃は、外側視床下部の刺激によって引き起こされるものに似ていました。
刺激が始まるとネコは鼻を地面に近づけ、毛を少し逆立てた状態で素早く動き、まっすぐラットに近づき、頭と首を繰り返し噛み、たいていは最初の数回の噛みつきでラットを殺しました
ネコはラットの尻尾、腹、背中を噛むこともありましたが、噛みつきは最終的に首と頭に向けられていました。
いくつかの試験では、ネコはラットを無視してテストケージの周りを数回旋回し、その後突然ラットに襲いかかり獰猛に噛みつきました。
この攻撃は、瞳孔が散大し、尾と背中の正中線に沿って毛が逆立ったこと以外、自律神経の活性化の兆候がありませんでした。
外側視床下部と腹側被蓋野は欲求回路の一部であると考えられています。
実際、外側視床下部や腹側被蓋野の刺激で、捕食攻撃だけでなく、摂食や飲水、交尾などの動機付け行動が生じることが確認されています。(4ページ目「快楽と欲求」参照)
扁桃体
またフリン博士は、捕食攻撃が扁桃体の外側核の電気刺激によって促進されることを発見しました。(Egger and Flynn 1963)
実験ではまず、外側視床下部を電気刺激して捕食攻撃を誘発しました。
これは噛みつきを特徴とする攻撃もあれば、前肢で叩く攻撃もありました。噛みつきと叩きの両方が混在する形態の攻撃もありました。
ここで扁桃体の外側核の電気刺激を加えると、この捕食攻撃が促進されました。
具体的には、ネコはより早く攻撃を仕掛けるようになりました。
後続の研究では、扁桃体の外側核の他、中心核も捕食攻撃を促進することが示されています。(Block et al. 1980)
一方で防衛的激怒は、扁桃体の内側核・基底内側核の電気刺激で促進されることが示されています。(Stoddard-Apter and MacDonnell 1980)
実験ではまず、視床下部の腹内側核を電気刺激して、防衛的激怒を誘発しました。
ネコはシャーと鳴き、毛を逆立て、瞳孔を開き、耳を後退させ、うなり声を上げました。
ここで扁桃体の内側核か基底内側核に電気刺激を加えると、この防衛的激怒が促進されました。
具体的には、ネコはより早くシャーと鳴くようになりました。
水道周囲灰白質
防衛劇激怒と捕食攻撃の区別は、中脳の水道周囲灰白質にも存在することがわかりました。(Bandler 1975, Bandler 1977)
水道周囲灰白質の背側部に電気刺激を与えると、ネコは激しい怒りを顕わにしました。
ネコは「シャー!」と鳴き、「カッ!」と怒声を浴びせ、唸り声を上げ、歯をむき出しにし、耳を後ろに倒し、背中を反らせ、前足で何度も叩きました。
一方、水道周囲灰白質の腹側部に電気刺激を与えると、ネコは捕食を開始しました。
ネコは鼻を地面に近づけて素早く移動し、他っとにまっすぐに向かい、首と頭を繰り返し噛み、たいていは最初の数回の噛みつきでラットを殺しました。
瞳孔の散大と毛の逆立ち以外には、自律神経の活性化の特徴をともないませんでした。
攻撃の脳地図
視床下部
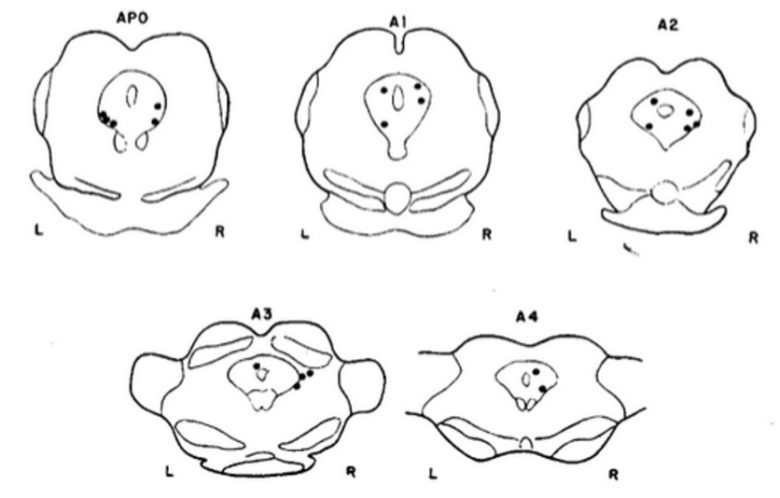
ニュージャージー医科大学のアラン・シーゲル博士もネコの二種類の攻撃行動を誘発する実験を行い、先行する研究を裏付けました。(Siegel and Brutus 1990)
博士の実験でも同様に、ネコの内側視床下部の電気刺激で防衛的激怒が誘発され、外側視床下部では捕食攻撃が誘発されました。
博士はフリン博士の研究を拡張し、より高い精度でネコの視床下部で攻撃が誘発される領域を調査し、詳細な脳地図を描きました。
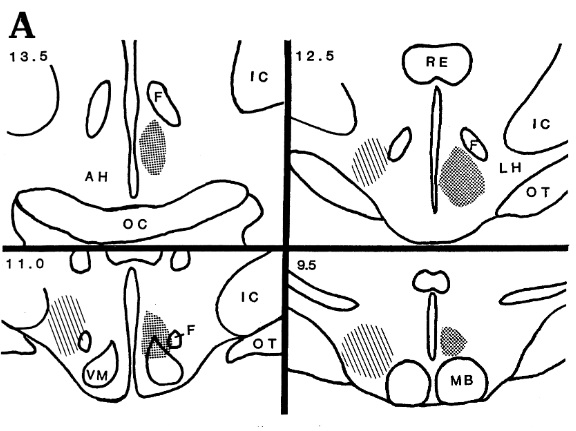
VM : 腹内側核、F:脳弓、MB:乳頭体、LH:外側視床下部、OT:視索、RE:視床連結核、IC:内包。

水道周囲灰白質
シーゲル博士は中脳の水道周囲灰白質についても、ネコの二種類の攻撃について先行する研究を裏付けました。(Shaikh et al. 1987)
博士の実験でも同様に、水道周囲灰白質の背側部の電気刺激で防衛的激怒が誘発され、腹側部では捕食攻撃が誘発されました。
博士は水道周囲灰白質についても、より高い精度でネコの攻撃が誘発される領域を調査し、詳細な脳地図を描きました。
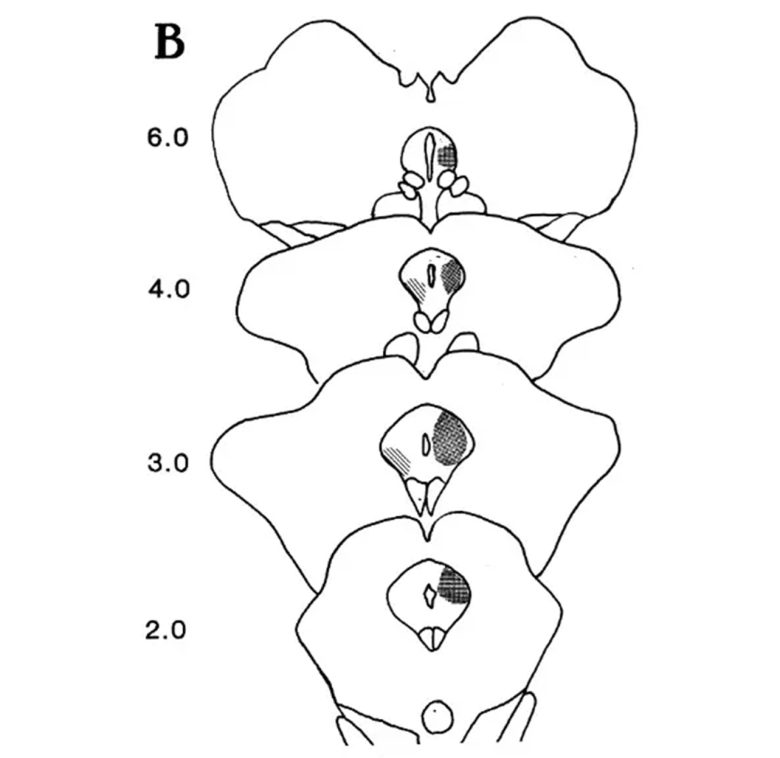
水道周囲灰白質の攻撃の脳地図。点描の領域 (背側部)が防御的激怒、縞々の領域 (腹側部) が捕食攻撃を誘発する。 (Siegel and Brutus 1990)
サル
サルを使った電気刺激による激怒・攻撃行動の誘発実験が、限られた研究グループによっていくつか行われています。
神経回路の探求というよりは、社会的な状況における行動の変化に着目した実験が多いです。
これらのサルを使った脳電気刺激の実験は、人間社会でのマインドコントロール疑惑を呼び起こしたと言われています。(Blackwell 2014)
デルガドの実験
イェール大学のホセ・デルガドは、コロニー内で生活するサルの脳に電気刺激を与え、社会的な状況での行動への影響を調査する実験を行いました。(Delgado 1964)
ボスザルの水道周囲灰白質への電気刺激は、即座に攻撃性の即座を引き起こしました。
ボスザルはケージ内を走り回り、群れの他の個体を攻撃し、追いかけ、逃げられなかった個体に噛みつきました。
この攻撃行動のパターンは自発的な攻撃と同じでしたが、より激しく執拗で、攻撃は刺激が与えられている限り続きました。
攻撃はよく組織化されて狙いが定まっており、社会的な選択を示していました。なぜなら、攻撃は特に、以前からボスが敵対的な関係にあったオスの4番のサルに向けられていたからです。
ボスの攻撃性の高まりは、ブランコを噛んだり、ケージを揺すったり、ケージの前面のガラスに映った自分の姿を攻撃したりすることでも現れました。
攻撃性の社会的な広がりもみられました。
ホスザルが電気刺激を受けるたびに、友好的で親密な関係にあったメスのサル7番がホスザルに加わり、群れの残りのサルを攻撃しました。
いくつかのケースでは、他のサルよりもずっと小さいにもかかわらず、7番のメスザルから攻撃を仕掛けました。



これらの友好的な関係と攻撃する動物の選択は、コロニー内の以前の関係によって決定され、過去の自発的な群れ形成と衝突を再現したものでした。 (Delgado 1967)
脳刺激は社会的な好き嫌いは変えず、攻撃行動の回数だけが増加しました。
したがって脳の電気刺激は攻撃における定型的な運動を誘発したのではなく、動物の感情状態を変化させたものとみなされました。

ボスザルは視床に電気刺激を受けると、3番のサルを100%の確率で威嚇した。写真はデルガドの別の研究から。(DELGADO 1966)
デルガドはまた、脳の電気刺激によって誘発される攻撃的な反応は、サルの社会的地位に依存していることも示しました。(Plotnik et al. 1971)
ただしこの実験で刺激された部位は視床や三叉神経などであり、おそらく痛みによって引き起こされた二次的な攻撃性であることに注意する必要があります。
たとえばサルのA6は、電気刺激後に支配的なサルを威嚇したり攻撃したりすることはなく、代わりに刺激後に顔をしかめるだけに終わりました。
一方で、A6は従属的なサルを威嚇・攻撃し、より頻繁にはおもちゃのトラを威嚇・攻撃しました。



ヤーキーズ霊長類研究所の実験
ヤーキーズ霊長類研究所でも、コロニー内で生活するサルの脳に電気刺激を与えて、社会的な状況での行動への影響を調査するという実験が行われました。(Robinson et al. 1969)
ある実験では、脳電気刺激によってサルのコロニーの地位を変化させることができることが示されました。
群れは支配的なオス、従属的なオス、そしてメスで構成されていました。支配的なオスはケージ内を歩き回り、従属的なオスを追い払うか積極的に攻撃し、メスを独占していました。
従属的なオスの視床下部に電気刺激を与えると、支配的なオスに対する攻撃が生じ、支配的なオスは激しく反撃しました。
メスや無生物に対する攻撃は行われませんでした。
攻撃は通常、刺激の開始後5秒以内に発生し、刺激の終了後に戦闘は停止しましたが、支配的なサルが戦いを続けることもありました。
これらの戦いは激しく凶暴で、裂傷や脱毛をもたらしました。
当初、従属的なサルはほとんどの戦いに敗れ、刺激の合間は従順な身振りをし続けました。
従属的なサルへの脳刺激が繰り返されるにつれて、支配的なサルはあまり効果的に戦わなくなり、刺激の合間は静かに座っているようになりました。
1つのグループでは10回目の刺激で、もう一方のグループでは43回目の刺激で、2頭のオスの地位が逆転しました。
この変化が近づくにつれて、最初は支配的だったオスは攻撃されるたびに大きな悲鳴をあげるようになりました。
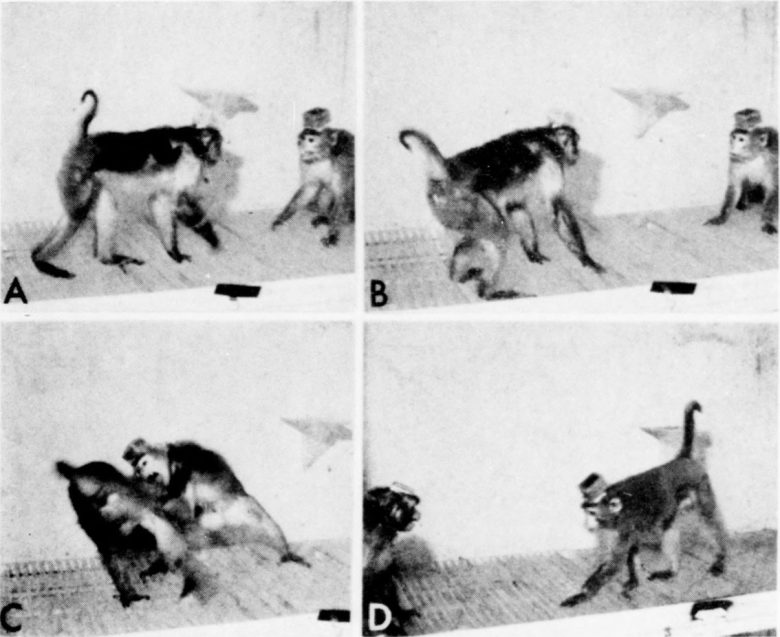
支配権の逆転後、実験サルに威嚇行動のみを引き起こす短時間の微弱な電気刺激を与えたところ、脅されたオスに恐怖の発声と逃走を引き起こすのに十分でした。
新たに優位に立ったオスは、ケージの中を自由に歩き回り、かつて優位だったオスを簡単に追い払い、コロニー内のメスにマウントし、やがてメスに毛繕いされるようになりました。
同研究センターの別の実験では、性別と優位性が、サルの誘発された攻撃反応に及ぼす影響が調査されました。(Alexander and Perachio 1973)
実験ではまず、サルの視床下部の電気刺激で攻撃反応が誘発されることを確認しました。
弱い電気刺激ではサルは威嚇だけを行い、刺激の強さが増すと攻撃へと変化しました。
攻撃が起こると、噛みつき、平手打ち、掴みなどの格闘戦が続きました。
続いて性別と優位性がサルの攻撃性に与える影響が調べられました。
支配的なサルと従属的なサルの両方が存在する状況では、実験サルは電気刺激を受けると、従属的なサルを優先的に攻撃しました。
支配的なサルしか存在しない状況では、実験サルは攻撃に失敗するか、あるいは攻撃頻度が低くなりました。
また、オスはメスより優先的に攻撃されました。
しかし、メスだけが存在する状況では、刺激を受けたオスはメスを容易く攻撃できることが実証されました。
その他の実験
チューリッヒ大学のロベルト・フンスペルガ博士は、ネコと同様、サルでも視床下部の腹内側核への電気刺激で攻撃反応が誘発されることを示しました。(Lipp and Hunsperger 1978)
実験ではマーモセットが使われ、攻撃反応は以下の要素で構成されていました。
- 「声による威嚇」:「ケケケ……」という連続した鳴き声
- 「短い攻撃」:時折、相手または無生物をつかむ、切りつける、または噛む
- 「激しい攻撃」:ある程度の距離 (少なくとも30 cm) を素早く移動し、その後、対象を継続的に切りつける、つかむ、噛む、または追いかける
- 「格闘」:2匹のサルが互いにつかみ合い、切りつける、噛む



その他の動物
その他の動物でも、脳電気刺激によって2種類の異なる攻撃反応が生じることが示されています。
オポッサム
オポッサムでも視床下部の異なる部位の電気刺激により、ラットに対して2種類の異なる攻撃反応が生じることが示されています。(Roberts et al. 1967)
内側視床下部の刺激では防御的な威嚇行動が誘発されました。
オポッサムは口を大きく開け、唸り声を上げ、頭と肩を左右に振り、後ずさりしました。
その後、後部が保護される角に後ずさり、開いた口を弧を描くように前方へ振り、威圧的な防御姿勢を取りました。
攻撃的な外見とは裏腹に、反応はほぼ純粋に防御的なものでした。
ぬいぐるみのオポッサム、生きたオポッサム、または口元近くで動かした棒で試してみると、実際の噛みつきは稀で短く、適切に方向づけられていませんでした。
外側視床下部の刺激では捕食攻撃が誘発されました。
攻撃パターンの主な要素は、捕獲、振り回し、殺し噛みつき、そして噛み砕きでした。
オポッサムはラットをじっと見つめ、首を伸ばして近づき始めました。
ラットを口の中に捕らえた後、オポッサムは激しく頭を振り回し、ラットに損傷を与えました。
頭を振り回す合間に、オポッサムはラットを殺すための噛みつきを数回繰り返しました。ラットが抵抗を続ける限り、この噛みつきは繰り返されました。
ラットが抵抗をやめると、頭を振り回す動作は止まり、噛みつきは頭部へと移りました。頭部への噛みつきはリズミカルになり、噛み砕くような音が聞こえました。
活発なラットは短時間であればオポッサムを撃退できましたが、刺激の強度が増すと、オポッサムの攻撃傾向は強まり、ラットの抵抗は無駄に終わりました。
攻撃は常に刺激終了後、即座に終了しました。
刺激により生後60~70日のオポッサムの赤ちゃんに対する捕食攻撃すら生じ、マウスのように無残にも食べられてしましました。
自発的な捕食攻撃は刺激によって引き起こされる攻撃と似ていましたが、より緩やかに始まり、それほど激しくなく、時には一時的な後退を伴うこともありました。
ラット
ラットでも同様に、視床下部の異なる部位の刺激により防衛的激怒と捕食攻撃という2種類の攻撃が生じることが確認されています。
視床下部の腹内側核およびその背外側の領域への電気刺激では、防衛的激怒が生じました。(Kruk et al. 1983)
攻撃行動は主にジャンプ攻撃と咬みつき攻撃で構成されていました。
ジャンプ攻撃では、刺激を受けた雄ラットは別の雄に近づき、その前方へ飛び上がりました。
ジャンプしながら相手の頭部を噛もうとすると同時に、たいていは直立している防御的な相手の腹部に激しい飛び込みを加えようとしました。
噛みつき攻撃では、刺激を受けたラットは飛び上がることなく、単純に相手の頭に噛みつきました。
どちらの攻撃の後にも、激しい格闘戦が続くことがありました。
外側視床下部の電気刺激では、マウスと幼若ラットに対する捕食攻撃が生じました。(Woodworth 1971)
捕食攻撃の主な要素は、捕獲、ジャンプ攻撃、そして殺し咬みつきでした。
前足で掴むのを伴う捕獲噛みつきは、体の前部に向けられることが多かったです。
捕獲噛みつきに次いでジャンプ攻撃が起こり、これは非常に速いため、ハイスピードフィルムでしか追跡できませんでした。
ジャンプしている間も標的は口と足で掴まれ続け、ラットは空中でも後ろ足で標的を蹴ることが多かったです。

殺し咬みつきは、攻撃対象の体中への反復的な咬みつきでした。
顎を閉じるたびに頭部が前方に突き出され、前足は攻撃対象を床に固定するか、口の中での位置を変えるために使用されました。
刺激が消失してから1秒以内に噛みつきは常に止まりましたが、標的がすぐに落とされるとは限りませんでした。
イヌ
イヌでも視床下部への電気刺激で激怒と攻撃行動が生じることが示されています。(Fonberg 1967)
刺激開始直後にイヌは歯をむき出しにし、目をぐるりと回し、激しく唸り声を上げ、唸ったり吠えたりした後、ハーネス、ひも、棒など、近くにある物体を攻撃し始めました。
実験者の手が露出していると、その手を噛もうとすることもありました。棒で脅かされると、それを歯で掴もうとしました。全体として、攻撃は適切に方向づけられていました。
刺激がなくなるとほぼすぐに、イヌは以前の穏やかで友好的な状態に戻り、触れられても攻撃性を示しませんでした。
異なる日に同一地点を刺激すると、同じ行動が再現されました。 これは変化なく、数週間、あるいは数か月間にわたって確認することができました。
ウサギ
ウサギの視床下部あるいは水道周囲灰白質を電気刺激すると、肢の踏みつけ (thumping)、いわゆる足ダンが誘発されました。(Black and Vanderwolf 1969)
刺激を受けたウサギは、両後肢を同時に上げ、地面に鋭く叩きつけ、特徴的な「ダン」という音を発しました。
この反応は通常、ウサギが動かずに立っていて、明らかに非常に警戒している時に起こりましたが、動いているときに観察されることもありました。
この行動には短いうなり声が伴うこともありました。最も一般的には、肢の踏みつけは刺激の停止から数秒以内に発生しました。
刺激を受けないウサギからは肢の踏みつけはほとんど観察されませんでした。
